
Фитогормоны виноградной лозы
Фитогормоны виноградной лозы
Фитогормоны виноградной лозы
Содержание материала
X. Дюринг
ФИТОГОРМОНЫ ВИНОГРАДНОЙ ЛОЗЫ
ВВЕДЕНИЕ
Понятие фитогормоны охватывает 5 групп фитогенных веществ, передвигающихся в растении и в незначительных дозах индуцирующих или контролирующих различные физиологические процессы. Обозначенные как “химические сигналы”, гормоны в большей части обладают высокой подвижностью во всем растении, так что регулирующие функции этих веществ проявляются в месте их синтеза или, в известной степени при коррелятивных отношениях, в других органах. В более узком смысле к фитогормонам причисляются ауксины, гиббереллины, цитокинины, абсциссины и этилен. В разделе, где описываются выявляемые аналитическим путем фитогормоны виноградной лозы, речь идет и о дополнительных веществах, оказывающих ингибирующее действие, которые использовались в биологическом тексте. В следующем разделе рассматриваются вопросы действия синтезируемых аналогов фитогормонов и аналогичных им веществ, напоминающих фитогенные гормоны.
Исследовательская работа в целях определения гормонов виноградной лозы и их действия в последние годы разрослась до такой степени, что в объеме столь небольшой работы не представляется возможности охватить новейшую литературу по этому вопросу примерно после середины 60-х годов. Данные по этому вопросу можно найти в следующих обзорах: А11еwе1dt с сотр. (1967, а, b, 1975, 1977), Coombe (1973, 1976а), During,Allewе1dt (1974), During (1976a), Hidalgo, Candella (1973), а также в диссертациях Lott (1969), Hifnу (1971), During (1972), Ju1iiard (1973), Waitz (1975), Luckert (1976), Vollmer (1976), Koch (1977).
ЛОКАЛИЗАЦИЯ ФИТОГОРМОНОВ
Фитогормоны в вегетативных органах
Ауксины. Сезонная ритмика фаз роста и покоя виноградной лозы дает основание считать, что стимулирующие и подавляющие рост вещества участвуют в регулировании роста в отдельные времена года. Исследования Spiegel (1954) и Мashеvа (1975) показывают, что вещества, проявляющие ауксинную активность в биологическом тексте с побегами и почками, доказуемы, причем наиболее высокое их содержание установлено до распускания почек, после полного исчезновения растворимой в эфире ингибирующей фракции.
Эти результаты подтверждаются газохроматографическими анализами I s о d а (1975). Lilov и Nikolova (1974) установили, что физиологически молодые части побега, как и находящиеся под ними листья, отличаются большим содержанием ауксинов по сравнению с более старыми зонами побегов и листьями под ними. В главных и боковых корнях содержание ауксинов в течение вегетационного периода повышается, а после наступления зимнего покоя понижается. Наблюдавшийся Masheva (1975) базипетальный транспорт ауксинов свидетельствует в пользу предположения, что эти гормоны во время вегетационного периода передвигаются в корнях (Li1оv, Niко1оvа, 1974).
Цитокинины. Активность цитокининов в виноградной лозе была впервые доказана Nitsch и Nitsch (1965) в пасоке, а впоследствии, при применении того же метода, и в листьях (J а к о, 1970, 1977). V о 11 m е г (1976) подробно исследовал годичный ритм цитокининов в вегетативных органах разных сортов Vitis vinifera и на различных подвоях в условиях теплицы и открытого грунта. Низкое содержание их в зимних глазках в фазу эндогенного покоя сопровождается повышением активности в период после покоя до прорастания почек. До наступления сокодвижения это повышение, очевидно, объясняется синтезом или активацией цитокининов в почках, после чего наступает дополнительное накопление цитокининов, вероятно, из корней. Максимальное содержание цитокининов, установленное во время прорастания почек, сопровождается снижением его в молодых побегах до минимальных значений через 4—6 недель после прорастания. В июне содержание цитокининов вновь повышается в более старых листьях, тогда как в побегах и зимних глазках их содержание уменьшается с наступлением осени. Наподобие листьев, и в корнях установлено увеличение цитокинина во время вегетационного периода; более молодые части корней, однако, богаче цитокинином, чем более старые. Конус нарастания побегов также содержит больше цитокинина, чем листья. У всех исследовавшихся сортов и сочетаний прививок установлено повышение концентрации цитокинина по мере усиления интенсивности сокодвижения до прорастания почек и понижение — с ослаблением интенсивности плача после распускания почек. Очевидно, концентрация цитокинина в пасоке определяется преимущественно привоем.
Исследования Lilov и Andonowa(1976) подтверждают эти результаты. Во время роста ягод наступает снижение содержания цитокининов в листьях, а во время их созревания оно вновь повышается. Примерно до июля в побегах и корнях устанавливается повышение цитокинина, после чего его активность начинает снижаться до середины ноября, а затем опять повышается в декабре. Итак, за исключением листьев, во время эндогенной фазы покоя во всех органах виноградной лозы устанавливается незначительная, а впоследствии в различной степени возрастающая активность цитокининов.
Исследования в связи с повышением содержания цитокининов при различной температуре показали, что хранение в условиях низкой температуры (+1°С) повышает активность цитокининов в вызревших побегах (Skene, 1972), а также, что разная температура питательного раствора приводит к изменению содержания отдельных цитокининовых фракций в корневом экссудате (Skene и Keridge, 1967). Нарушение соотношения источник — потребитель (source — sink) у плодоносящих кустов путем удаления гроздей понижает активность фотосинтеза и в то же время усиливает образование цитокининовых глюкозидов в листьях (Hoad с сотр., 1977). Повышение содержания цитокинина примерно в 20 раз установлено в пасоке, когда в питательный раствор добавляли хлорхолинхлорид (ССС) (Skene, 1968, 1970).
Наоборот, снижение активности цитокинина в корнях отмечалось при уменьшении доз N или К. В условиях без удобрения NPK в корнях не обнаруживалось цитокининов (Jако, 1974).
Гиббереллины. До прорастания почек Isоdа (1973) установил повышение активности гиббереллинов (GA) в почках. В корневом экссудате и в экстрактах из листьев и корней Skene (1967) обнаружил вещества, Rf- значения и биологическая активность которых соответствовали GA4 и GA7. В усиках установлено большее содержание гиббереллина, чем в других органах побега; закручивание усиков сопровождается ростом гиббереллинов (Маnanкоv, 1976). По мнению Kocherzhenko и Maiko (1967), между содержанием гиббереллина в листьях и ростом существует тесная корреляция. Так, например, в условиях продолжительного освещения и длинного дня в листьях быстрорастущих растений при помощи теста прорастания гороха установлены гиббереллины, которых не обнаружено в условиях короткого дня при редуцированном росте. Удаление гроздей, по данным Hoad с сотр. (1977), ведет к уменьшению гиббереллиноподобных веществ в листьях.
Абсциссины и ингибиторы. Впервые в пасоке виноградной лозы при помощи биологического теста Rapp и Ziegler (1971)установили абсциссиновую кислоту на основе Rf-значений в различных жидкостях, поведения в ультрафиолетовой области, а также инфракрасного и массового спектров. На основе этих методов впоследствии были исследованы сезонные изменения абсциссиновой кислоты в побегах и листьях: содержание абсциссиновой кислоты в листьях, где она синтезируется, не претерпевает существенных изменений в течение вегетационного периода. В побегах, наоборот, обнаруживается четкая сезонная динамика, причем максимум устанавливается в эндогенную фазу покоя (During, Alleweldt, 1973; Cheng с сотр., 1974). Решающим фактором повышения содержания абсциссиновой кислоты можно считать уменьшение длины дня, что, однако, не снижало ее количества в листьях, подвергнутых раздражению фотопериодом. Взамен этого, уменьшение длины дня увеличивает абсциссиновую кислоту в побегах, что свидетельствует о ее передвижении из листьев к побегам (Alleweldt, During, 1972). В апикальных частях побегов нередко устанавливалось большее содержание абсциссиновой кислоты, чем в секторах у основания (Lilov, Angelova, 1977а; During, неопубликованные данные). Как показывают опыты Lоveys и Kriedemann (1973), наблюдается быстрая и четкая реакция содержания абсциссиновой кислоты в листьях на продолжительный водный дефицит; при пониженной интенсивности транспирации и фотосинтеза, а также при повышении водного потенциала, содержание абсциссиновой кислоты в листьях увеличивается, тогда как после повторного полива наблюдается обратное. Наряду с повышением содержания абсциссиновой кислоты в листьях увядающих виноградных кустов устанавливается еще большее его повышение в побегах (During, Sсiеnz а, 1975), что, по всей вероятности, приводит к прекращению роста побегов в длину. Исследования показали, что суточный ход транспирации в большинстве случаев находится в отрицательной корреляции с содержанием абсциссиновой кислоты в листьях (During, Scienza, 1975). Из листьев выделена также близкая абсциссиновой фазеиновая кислота (Loveys, Kriedemann, 1974).
В нейтральных, основных и кислых экстрактах, из побегов Тiziо с сотр. (1968) установили при помощи биологического теста ингибиторы. Weaver с сотр. (1968) и Sarkisova с сотр. (1976) установили в начале периода покоя повышение феноловых ингибиторов в побегах и почках, содержание которых уменьшается с началом вегетации. Это эндогенное содержание ингибиторов непосредственно связано с морозостойкостью отдельных сортов (Sarkisova с сотр., 1976).
Фитогормоны в репродуктивных органах
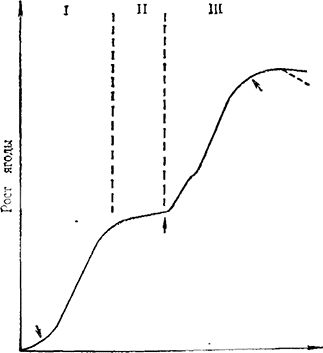
Рис. 1. Динамика содержания ауксинов во время роста ягоды винограда
I — цветение; II — начало созревания; III — физиологическая зрелость
Ауксины. Впервые ауксиноподобные вещества в ягодах винограда установили Nitsсh с сотр. (1960) и Сооrubе (1960). При помощи теста A vena mesocotyle установлено повышение содержания ауксиноподобных веществ в I фазе роста ягод (рис. 1) и понижение его при временном приостановлении роста во II фазе. Исследования Alleweldt и Hifny (1972) и Farmahan и Pandey (1976) подтверждают этот ход ауксиноподобных веществ.
Ауксин в незначительной концентрации доказан в ягодах при помощи газовой хроматографии (Сооmbе, Hale, 1973) и хроматографии с использованием жидкости под большим давлением (During, 1977). Применение гиббереллина калия повысило уже через 3 h после обработки активность ауксинов в ягодах; этот повышенный уровень ауксинов вновь понижается во II фазе роста ягод (Bertrand, Weaver, 1972а). Во время стратификации активность ауксинов в семенах повышается, достигая максимума при прорастании (Сhоhan, Dhi11оn, 1976; Касhru с сотр., 1969).
Цитокинины. Новейшие исследования при помощи теста в каллюсе сои показывают, что неидентифицированные до сих пор активные вещества (Waitz, 1975; Сhаско с сотр., 1976; Lilov, Andonova, 1976) появляются в ягодах преимущественно в начале их роста. Относительно содержания цитокининов в отдельные фазы роста имеется немного данных, которые, однако, совпадают. Waitz (1975) установил совсем незначительные концентрации цитокинина в соцветиях в полном цветении, в то время как Lilov и Andonova в этой фазе обнаружили большие концентрации, чем в развивающихся впоследствии ягодах. Пробы в исследованиях брали в один, соответственно в два срока. В разгар цветения и во время образования ягод Lilov и Andonova обнаружили незначительную концентрацию цитокининов, а и Waitz сообщает о цитокининовом минимуме. В противовес этому Сhаско с сотр. (1976) установили максимальные значения, уменьшающиеся в фазу интенсивного роста (I фазу).
Если пренебречь наблюдаемым в III фазе повторным повышением содержания цитокинина, то можно принять, что сделанные констатации о появлении цитокининов в ягодах преимущественно в первую фазу усиленного роста в значительной мере совпадают.
Гиббереллины. Наряду с ауксинами Сооmbе (1960) впервые установил гиббереллиновую активность в незрелых ягодах многих сортов. С началом нарастания ягод Iwahоri с сотр. (1968), Itо с сотр. (1969), а также Lilον и Christov (1977) обнаружили максимальное содержание гиббереллина, задерживающие на одном уровне около двух недель, а затем понижающееся. До сих пор повышение содержания гиббереллиноподобных веществ в III фазе созревания было установлено только Farmahan и Pandey (1976). Бессемянные сорта почти без исключения отличаются меньшей активностью гиббереллина по сравнению с сортами с семенами (Сhai1akhуаn, Sarkisova, 1965; Weaver, Pool, 1965а,b; Lilov, Christov, 1977; Nazemi11e, 1977), что выражается несколько замедленным нарастанием незрелых ягод с семенами (Iwahori с сотр., 1968). Незначительное содержание гиббереллина в незрелых ягодах соответствует его меньшим значениям также в листьях, соцветиях и гребнях, тогда как у сортов с семенами в генеративных органах установлено большое содержание гиббереллина, чем в вегетативных (С hailakhyan, Sarkisova, 1965). Weaver и Pool (1965а) обнаружили активность гиббереллиноподобных веществ до оплодотворения в узлах, несущих соцветия. Кольцевание вызывает изменения в спектре гиббереллинов, что означает, что содержание гиббереллина в кислой фракции повысилось, а в нейтральной — понизилось (Weaver, Pool, 1965 b, с).
Абсциссины. О локализации абсциссиновой кислоты в ягодах винограда сообщают Coombe и Hale (1973), Inаbа с сотр. (1976) и During (1973, 1974, 1977), которым при помощи газовой хроматографии, соответственно хроматографии в жидкости под высоким давлением, удалось установить увеличение этого гормона с началом созревания ягод и понижение — перед окончательным созреванием. During (1974) установил почти параллельное повышение абсциссиновой кислоты в мякоти ягод и семенах, в то время как Coombe и Hale (1973) не удалось установить увеличения абсциссиновой кислоты в семенах; Lilov и Angelova (1977b) и Scienza (личные сообщения) также обнаружили лишь незначительное количество абсциссиновой кислоты в семенах. Lott (1968, 1969), однако, установил в выделенных семенах винограда высокое содержание абсциссиновой кислоты, которое уменьшается во время стратификации параллельно повышению степени их готовности к прорастанию. Констатации (неопубликованные) During и данные Chоhan и Dhil1 on (1976) подтверждают эту зависимость. По мнению Coombe (1976a,b) содержание абсциссиновой кислоты повышается примерно в течение 10 недель после цветения, причем в кожице ягод увеличение наступает раньше и быстрее, чем в мякоти.
Этилен. Подобно неклимактерическим плодам черешни (не претерпевающим резких изменений в дыхании во время созревания), в ягодах винограда были установлены совсем незначительные концентрации этилена, во время созревания проявляющие тенденцию к снижению (Coombe, На1е, 1973). В новейших исследованиях Inаbа с сотр. (1976), Косh (1977) и А11еwе1dt и Koch (1977) также установлено внутриклеточное содержание этилена. Независимо от относительного значения (на ягоду или грамм свежего веса), в начале II фазы установлено повышение содержания этилена, которое, однако, перед началом созревания вновь быстро понижается.
Фитогормоны виноградной лозы – Действие фитогормонов винограда и аналогичных им веществ
Содержание материала
Вегетативные органы
Ауксины и аналоги ауксинов. После обнадеживающих опытов Мu11еr- Stо11 (1939) в связи с воздействием ауксином на корнеобразование и развитие побегов виноградной лозы в целях производства привитых укорененных саженцев вновь были выдвинуты вопросы о корнеобразовании и аффинитете. Согласно исследованиям Тiziо (1967), даже незначительные концентрации индол-3-масляной кислоты (0,25—2,5 ppm) стимулируют корнеобразование легко окореняющегося сорта Мальбек, тогда как для труднее окореняющегося подвоя Кобера 5 ББ необходимы 0,25—25 ppm. В начале вегетационного периода, вследствие высокого содержания ингибиторов в побегах, возникает потребность в повышенных концентрациях ауксина; с увеличением эндогенных ростовых веществ и повышением способности к каллюсообразованию и корнеобразованию концентрацию применяемых ауксинов можно снизить (Тiziо, 1967). С целью улучшения окоренения зеленых черенков Turetskaya c сотр. (1966) предлагают 14-часовое погружение в 150 ppm индол-3-масляной кислоты. Это повышает процент окоренившихся черенков сорта Мадлен Анжевин с 70 до 100, а трудно окореняющегося сорта Тихоновский — с 4 до 74. По-видимому, генетически обусловленное более легкое окоренение некоторых сортов основывается на ускоренном и более интенсивном синтезе и активировании индол-3-уксусной кислоты у этих сортов (Turetskayac сотр., 1966). По мнению Eccher и Маrrо (1971), оптимальная концентрация наиболее часто используемых на практике растворов нафтил-3-уксусной кислоты зависит главным образом от величины черенков. Так, например, у длинных черенков высокие дозы (до 300 ppm нафтил-3-уксусной кислоты) могут стимулировать корнеобразование, тогда как небольшие дозы оказывают токсическое действие. Для видов Р. tricuspidata и Р. guinguefolia оптимальной для корнеобразования является концентрация 10- 6 до 10-4 (Lеrоuх, 1965). Shumann и Uhl (1975) усматривают в использовании метилэфира нафтил-3-уксусной кислоты и других стимулирующих рост и корнеобразование веществ технические преимущества для производства саженцев винограда. По-видимому, индуцированный индол-3- уксусной кислотой Rhizokalin (Ju11iаrd, 1966) в своем базипетальном передвижении не нуждается в мобилизации через посредство базипетально передвигающейся индол-3-уксусной кислоты (Ju11iаrd, 1967). По длине побегов создается корнеобразующий градиент, который не изменяется экзогенно под влиянием питательных и ростовых веществ. Очевидно, вместе с индол-3-масляной кислотой и гиббереллин (10-7—10-8М) оказывает стимулирующее воздействие на корнеобразование (Ju11iаrd, 1970а).
Погружение подвоев (V. solonis х V. Riparia 1616) на 24 h в концентрацию 50 ppm нафтил-3-уксусной кислоты непосредственно перед прививкой сорта Султанина определяет наибольший выход привитых саженцев, тогда как индол-3-уксусная кислота, индол-3-масляная кислота и гиббереллин оказали более слабое воздействие (Stinо с сотр., 1977). Если нижнюю треть подвоев прививок погрузить на 12 h в раствор ростовых веществ (0,1 % индол- 3-уксусной кислоты, 0,05% индол-3-масляной кислоты или 0,006% а-нафтил- 3-уксусной кислоты), повышается содержание питательных веществ в привое, а 90—95% привитых кустов вступают в плодоношение на третий год после посадки. Плодоносность глазков повышается, повышается и урожай винограда в первый год плодоношения на 7—92% (14—25 c/ha против 13 с/ha в контроле). Средний вес гроздей, однако, понижается (Khanin, Stepanova, 1970). По всей вероятности, лучшее срастание объясняется влиянием ауксинов на каллюсообразование (Fallot, 1970).
Как показывают опыты Sarkisova и Chailakhyan (1968) и Skene (1971), ауксин, по-видимому, участвует в метаболизме углеводов в черенках. Так, индол-3-уксусная кислота вызывает увеличение содержания сахаров и снижение концентрации крахмала (Skene, 1971). β-Индол-3-масляная кислота стимулирует физиологическую поляризацию черенков посредством быстрого смещения запасных веществ в сторону основания, где начинается их использование на корнеобразование; наконец, активизируется и ускоряется рост побегов (Sarkisova, Chailakhyan, 1968). Ессhеr и Маrrо (1971) и Skene также наблюдали стимулирование обмена веществ, повышенную активность камбия и повышенное потребление углеводов после обработки ауксином. Однако, наряду со стимулирующими рост свойствами, ауксины обладают и ингибирующим действием. Так, например, 200—400 ppm нафтил-3-уксусной кислоты задерживают прорастание почек примерно на 5 дней (Di Cesare, 1968), а обработка индол-3-уксусной кислотой черенков сорта Мерло, нарезанных незадолго до прорастания почек, замедляет этот процесс, если обрабатывается конус нарастания (Liuni, Poppi Struma, 1973a, b). Индол-3-масляная кислота также задерживает прорастание почек примерно на 7 дней (Sarkisova, Chailakhyan, 1968). Ауксины подавляют прорастание зимних почек не только непосредственно, а и косвенно. Исследования Alleweldt и Istar (1969) показывают, что 500, соответственно 1000 ppm индол-3-уксусной кислоты сокращают рост в длину пасынков на 38—56% у подвергнутых вершкованию растений, т.е. при устранении доминирующего влияния конуса нарастания.
Цитокинины. В отличие от ауксинов, применение кининов (1000 ppm бензиладенина) вызывает преждевременный выход почек из состояния покоя (W е a v е г с сотр., 1974а,Ь), что, по-видимому, обусловливается ускоренным распадом ингибиторов, наблюдаемым после применения бензиладенина (Wеavеr с сотр., 1968). Только у неокоренившихся черенков кинины (прежде всего рибозиды бензиладенина) стимулируют рост побегов. По мнению Pool и Powell (1975), это свидетельствует об участии образовавшихся в корнях цитокининов в развитии побегов. Редуцированный под влиянием индол-3-уксусной кислоты рост пасынков в длину отчасти восстанавливается при добавлении кинетина (100 и 1000 ppm) (Alleweldt, Istar, 1969). Очевидно, и результаты, полученные Scuflaire и Brebion (1967), объясняются задерживающим онтогенез действием цитокининов. С этой целью авторы погружали виноградные листья с черешками на 8 дней в раствор кинетина (10-2, 10-5, 10-7 М) в благоприятных условиях (освещение, температура, влажность воздуха). В этих листьях установлена стабилизация обмена веществ. Накопление продуктов фотосинтеза в контрольных листьях, как и распад существующих уже веществ, ограничивались под влиянием кинетина. Применение бензиладенина стимулирует движение ассимилятов из одного листа к другому, а также накопление 14С в физиологически более старых листьях в условиях затемнения (Quinlan, Weaver, 1969).
Гиббереллины. Многочисленные работы Alleweldt (1964, 1977; Alleweldt с сотр., 1972,1977) показывают, что рост побегов в длину подвержен влиянию в первую очередь гиббереллина, причем сорта V. vinifera заметно чувствительнее, чем американские. Вызванное фотопериодически коротким днем подавление роста побегов в длину преодолевается при помощи гиббереллина, а не в результате индуцированного коротким днем покоя почек (Аllеwе1dt, 1964). Наблюдавшееся подавление прорастания почек гиббереллином (Alleweldt, 1961) подтверждает Еris (1976 a,b). Применение гиббереллина подавляет прорастание почек во время глубокого вынужденного покоя (Е г i s, 1976а), следовательно, зимний покой становится более глубоким (Julliard, Balthazar d, 1965). Наряду с ростом побегов, обработка гиббереллином (1—100 ppm) уменьшает диаметр корней (Skene, Mullins, 1967). Подобно кининам, очевидно, и гиббереллины регулируют движение ассимилятов из одного побега к другому (Quin1аn, Мu11ins, 1970). Гиббереллин (10 ppm) поглощается как листьями, так и междоузлиями и передвигается к верхушке побега. Лучшие результаты в отношении поглощения гиббереллина получены на поверхности молодых листьев у их основания (Wеavеr и др., 1966а).
Абсциссины. Подобно индол-3-уксусной кислоте, абсциссиновая кислота подавляет прорастание зимних почек (Julliard, 1970b; Liuni, Poppi Struma, 1973a). Обработка базальных и апикальных почек дала одинаковые результаты, тогда как индол-3-уксусная кислота действует только на апикальные (Liuni, Poppi Struma, 1973b). Увеличение содержания абсциссиновой кислоты в зимних почках в фазе глубокого покоя приводит к выводу, что налицо специфическое участие абсциссиновой кислоты в эндогенном глубоком покое почек (During, Bachmann, 1975; During, Kisma1i, 1975). Независимо от возраста листьев, под влиянием 20 μΜ абсциссиновой кислоты повышается содержание антоцианов в листьях (Рiriе, Mullins, 1976). Обработка абсциссиновой кислотой пластинок молодых листьев стимулирует развитие соцветий и предотвращает отмирание зародышевых цветоножек. Через 24 h после обработки устанавливается известное подавляющее воздействие абсциссиновой кислоты на включение аминокислот в протеин (Mullins, Osborne, 1970). Инфильтрация абсциссиновой кислоты оказывает быстрое действие на срезанные листья: всего спустя 20 min, в зависимости от введенной концентрации, отмечается закрывание устьичных щелей, достигающее максимума через 70 min, после чего начинается частичное и повторное открывание (During, 1976b). Это свидетельствует о значительной функции абсциссиновой кислоты в регулировании движения устьиц. В противовес этому, абсциссиновая кислота не участвует в вызванном сменой света и темноты движении устьичных щелей, ибо ее содержание остается неизменившимся до и после включения, соответственно исключения света (During, 1976 а,b). Фотонастическое движение устьиц в значительно большей степени должно было бы определяться изменением межклеточной концентрации СО2, повышение которой в виноградных листьях ведет к закрыванию устьичных щелей, а не к изменению содержания абсциссиновой кислоты в листьях (Lоvеуs с сотр., 1973). Наряду с недостатком воды и кольцевание побегов приводит к повышению содержания абсциссиновой и фазеиновой кислот при условии, что отток ассимилятов из листьев понижен или полностью прекращен (Loveys, Kriedemann, 1974). Такое повышение содержания абсциссиновой кислоты, вызванное обогащением ассимилятами, приводит к закрыванию устьичных щелей, а при недостатке воды — и к улучшению водного потенциала листьев (During, 1978). Близкая к абсциссиновой фазеиновая кислота определяется как специфический ингибитор фотосинтеза in vivo (Kriedemann с сотр., 1975).
Аналоги этилена. Обработка этефоном (400 ppm) подавляет апикальный рост побегов в длину и задерживает прорастание почек примерно на 8—10 недель. При этом действие этефона значительно продолжительнее и равномернее, чем влияние повторного прищипывания (Lavее и др., 1977). Weaver и Роо1 (1971а) также установили после обработки 3000 ppm этефона замедленное прорастание почек на следующий год, а при обработке во время цветения сводилась на нет апикальная доминантность, т.е. начиналось образование пасынков и разделительной ткани на апикальных узлах.
Виноградные стимуляторы
Стимуляторы роста приобретают все большую популярность. И дело не только в том, что они способствуют росту урожайности – они обеспечивают повышенное качество наших овощей и фруктов.
Большинство из биологически активных веществ в низких и очень низких концентрациях играют роль стимуляторов роста, способствуют повышению иммунитета, активизируют плодоношение. В высоких концентрациях эти же препараты оказывают действия, угнетающие физиологические процессы в растении. Так что лучше немного недодать, чем передать.
Гиббереллин
Оказывает влияние на работу хромосомного аппарата. В значительной степени влияет на длину стебля. Не влияет на рост корня, а в повышенных концентрациях в водной культуре даже ухудшает состояние корней. Известно, что спящие почки выводятся из состояния покоя действием пониженных температур. Температурный фактор в этих случаях может быть заменен обработкой гиббереллином.
Гиббереллин используют для повышения урожайности кишмишных (бессемянных) сортов винограда, характеризующихся сравнительно мелкими ягодами. Опрыскивание виноградной лозы раствором (30 мг на 10 м2) во время цветения или через 5-7 дней после цветения способствует увеличению размера ягод в 1,5-2,5 раза и повышению урожайности на 50-100%. К тому же на несколько дней ускоряется созревание винограда.
Положительно действует гиббереллин и на некоторые семенные сорта винограда: увеличивается количество ягод, возрастает малосемянность и бессемянность, разрыхляется кисть (что снижает поражаемость ягод фитопатогенами), ускоряется созревание. Не нашли гиббереллин? Покупайте ЗАВЯЗЬ, это то же самое, только с примесями. БУТОН – Это та же гибберленовая кислота, натриевые соли, микро- и макроэлементы. Если не можем изобрести ничего нового – тогда придумываем новые названия.
Гтерероауксин и корневин
Относятся к ауксиновой группе препаратов, которые стимулируют корнеобразование. Корневую систему растений перед посадкой замачивают на 8-24 часа в водном растворе гетероауксина. После посадки полезно полить растение этим же раствором. Для повышения эффективности действия гетероауксина при укоренении черенков проводят совместную обработку растений цирконом (1 ампула на 1 л воды) и гетероауксином (200 мг на 1 л воды). Для лучшей приживаемости корневую систему растений перед посадкой опудривают корневином.
Эпин
Антистрессовый гормональный препарат. Его гормоны поддерживают в норме иммунную систему растений, особенно в стрессовых ситуациях: пониженных температурах, заморозках, затоплениях, засухе, болезнях, действиях пестицидов, засолении почвы и т. д. К тому же растения становятся более устойчивыми к изменениям погоды. Важно и то, что выращенная продукция отличается высоким качеством и пониженным содержанием тяжелых металлов, нитратов, остаточных пестицидов.
Установлено также, что препарат обладает активизирующим влиянием на побегообразование винограда, плодово-ягодных культур, цветов, декоративных кустарников.
Биостим
Регулятор роста природного происхождения, полученный вытяжкой из растений. Представляет собой комплекс фитогормонов ауксиновой и гиббереллиновой природы. Биостим рекомендуется применять для улучшения укоренения саженцев. Препарат укрепляет иммунную систему растения и помогает ему пережить неблагоприятные периоды жизни, справиться с заболеванием. Опрыскивание растений биостимом способствует ускорению роста и повышению урожайности растений.
Иммуноцитофит
Препарат ауксиновой группы, способствующий значительному повышению иммунитета растений и их сопротивляемости многим распространенным заболеваниям: фитофторозу, мучнистой росе, бактериозам, серой гнили, милдью и т. д. Стимулирует ростовые процессы.
После обработки повышенная сопротивляемость болезням сохраняется в течение одного-двух месяцев. Иммуноцитофит предназначен для обработки винограда, плодовых и ягодных культур, картофеля, томатов, огурцов, капусты, лука, цветов.
Янтарная кислота
Стимулирует плодообразование. Опрыскивание растений 0,01% раствором янтарной кислоты ускоряет зацветание, растения меньше поражаются фитофторозом, урожай значительно увеличивается. Используют янтарную кислоту и для обработки томатов. Опрыскивание растений для повышения продуктивности проводят в период бутонизации (40-60мг/л) и повторяют трижды. Интервал между обработками – 7 дней. Расход раствора 2 литра на сотку.
Циркон
Регулятор роста и развития растений, корнеобразователь, индуктор цветения. Получен из растительного сырья. Повышает всхожесть и энергию прорастания семян. Ускоряет цветение, рост и развитие растений на 5-10 дней. Сроки созревания сокращаются на 1-2 недели. Урожайность возрастает на 35-60%. Применение циркона резко снижает степень поражения заболеваниями. Обладает высокой корнеобразующей активностью. Способствует укоренению черенков. Ускоряет их приживаемость при пересадках и снижает накопление тяжелых металлов.
Марс-У
Препарат комплексного действия – пленкообразующий стимулятор роста растений, содержащий биологически активные вещества природного происхождения и водорастворимые полимеры мягкого действия. Обеспечивает полную реализацию потенциальных возможностей роста растений, повышает устойчивость растений к экстремальным (климатическим и экологическим) условиям.
Риверм
Универсальное удобрение – стимулятор, в состав которого входит большое количество питательных веществ (микро- и макроэлементов), а также живые микроорганизмы (азото- и фосфобактеры). Не содержит никаких синтетических компонентов. Применение Риверма обеспечивает устойчивость растений к засухе и заморозкам. Также Риверм эффективно совмещать со средствами защиты растений, это значительно понижает их стрессовое состояние.
Но нельзя забывать – чудес на этом свете не бывает, и все стимуляторы, подкормки и прочие «полироли» показывают себя только на фоне высокой агротехники! Недостаточно просто «присесть на эту иглу», надо ее еще подкрепить хорошей кормежкой и правильными поливами.
Источники:
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/fitogormony-vinogradnoy-lozy.html
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/fitogormony-vinogradnoy-lozy-2.html
http://grapes.hozvo.ru/Vinogradnyiestimulyatoryi-9729




