
Более подробные данные о морфогенезе почек и соцветий – Формирование почек и соцветий
Более подробные данные о морфогенезе почек и соцветий – Формирование почек и соцветий
Плодовый сад
Страницы
Образование и развитие цветковых почек у косточковых культур
У косточковых культур одни почки закладываются как вегетативные, другие как цветковые.
Как уже указывалось, в практике плодоводства почки косточковых культур называют чистыми на основании их функционального назначения. Однако понятие «чистые» необходимо понимать условно, ибо внутри чистых цветковых почек находятся зачатки цветков, которые, как правило, не развиваются в функционирующие половые органы.
Только изредка можно наблюдать случаи, когда цветковые почки вишни развиваются так же, как и генеративные почки яблони. Тогда внутри них закладываются как вегетативные, так и цветковые части.
Сроки дифференциации цветковых почек вишни зависят от метеорологических условий весенне-летнего периода, состояния растения, условий питания, сортовых особенностей и других причин. Холодная и затяжная весна и сырое лето замедляют начало дифференциации, теплая весна и жаркое лето ускоряют этот биологический процесс. Сроки начала и окончания дифференциации почек у разных сортов одной и той же породы колеблются (12—15 дней).
Ко времени прекращения роста побегов закладывается верхушечная — ростовая почка. Формирование цветковых почек у вишни начинается с образования почечных чешуй и недоразвитых листьев. Затем происходит дифференциация конуса нарастания и образования цветочных бугорков.

Фазы развития цветковой почки вишни
1 — спящие цветковые почки; 2 — обособление цветочных бугорков; 3, 4 — начало развития внутренних органов цветка; 5 — формирование тычинок и пестичных бугорков; 6 — образование венчика; 7 — цветковая почка в разрезе перед уходом в зиму; 8 — раскрытый цветок; 9 — развитие тычинок и пестиков; 9а — пестик; 9б — тычинка; 10 — тетрады; 10а — пыльца; 11 — плодовая почка раскрытая; 12 — цветок раскрытый; 13 — цветение; 14 — цветок в разрезе; 14а — тычинка; 14б — пестик; 15 — плоды; 16,16а — почка и цветок после повреждения морозами
Дифференциация цветковых почек начинается в нижней части конуса нарастания с образованием валика, имеющего неясные очертания. Через шесть-семь дней на нем становятся отчетливо видны зачатки пяти цветочных бугорков.
Продолжая расти, они вытягиваются в столбики, и через 15—20 дней появляются зачатки чашелистиков. Через 30 дней после начала дифференциации формируются зачатки лепестков и тычиночные бугорки. Последним формируется пестик.
Рост лепестков венчика, чашечки, дифференциация тычинок и пестика проходят в течение всего августа и сентября. В зиму цветковые почки уходят с дифференцированными, но не с закончившими свое развитие частями цветка. Полностью цветок развивается и цветет в следующий вегетационный период, т. е. весной.
Таким образом, на развитие цветковых почек вишни уходит летне-осеннее время — с конца июля до начала августа (2—2,5 месяца) и ранняя весна — с половины апреля до цветения. Летне-осенний цикл развития продолжительнее, чем весенний. Весной развитие почек длится 25—30 дней. В это время растут органы цветка, формируются семяпочки и пыльца.
Цветение начинается в условиях Московской области в конце мая. Зимой вишня находится в состоянии покоя, в течение которого оттепели чередуются с морозами и иногда вызывают гибель цветковых почек и потерю урожая.
Сроки закладки и дифференциации цветковых почек зависят не только от погодных условий и ростовых процессов, но и других причин. Имеет значение географическое местоположение, высота над уровнем моря, широта, долгота местности, возраст дерева, его сортовые особенности. Так, с увеличением высоты над уровнем моря почки закладываются позднее, и наоборот. В северных районах почки закладываются позднее, чем в южных.
В одних и тех же районах произрастания цветковые почки семечковых пород более зимостойки, чем у косточковых. Наиболее часты случаи подмерзания и вымерзания почек на юге у абрикоса, персика, вишни, сливы. Отношение цветковых почек к низким температурам определяется степенью их развития и подготовленностью к зиме. В условиях Средней Азии теплые зимы с периодическими небольшими морозами понижают морозостойкость почек у абрикоса. Наоборот, умеренно холодные зимы с одинаково устойчивыми морозами менее губительны для цветковых почек. В Ташкенте, где зима холоднее и устойчивее, чем в южных районах Узбекистана, почки сохраняются лучше, чем в районе Термеза, где они зимуют в более поздней фазе развития.
Цветковые, почки персика, так же как и других косточковых культур, реагируют на низкие температуры в зависимости от степени их дифференциации, возраста дерева и закладки. По данным И. М. Рядновой цветковые почки, закладывающиеся в условиях Краснодарского края в сухую и жаркую погоду, быстро проходят фазы развития, рано выходят из периода покоя и весною часто погибают от весенних заморозков и незначительных морозов. Закладывающиеся в условиях прохладной погоды почки более зимостойки и менее подвержены действию возвратных холодов. Зимостойкость почек персика, по данным того же автора, зависит от возраста дерева и ряда агротехнических приемов.
На старых деревьях с ослабленным ростом цветковых почек закладывается много. Из периода покоя они выходят почти одновременно и в случае возвратных заморозков погибают. У сильно растущих деревьев в поре полного плодоношения цветковые почки закладываются неодновременно. Длительность их развития благоприятствует дифференциации в разное время и выходу из периода покоя также в разные сроки. В случае наступления возвратных заморозков погибает лишь часть почек, заложившихся в наиболее ранние сроки. Улучшение агротехники и усиленное питание, а также обрезка, увеличивающая рост и продляющая сроки закладки цветковых почек, способствуют повышению их зимостойкости.
Цветки плодовых и ягодных пород отличаются друг от друга временем созревания тычинок и пестиков, характером развитая, строением завязи и т. п. Одновременное созревание в цветке тычинок и пестиков носит название гомогинии.
Если в цветке раньше развиваются тычинки и пыльца, а позднее — пестики (например, у земляники, малины), такое явление называют протерандрией. При более раннем по сравнению с тычинками созревании пестика это явление носит название протерогинии. Оно наблюдается у некоторых сортов яблони, груши, сливы.
Завязь бывает верхняя, средняя и нижняя. Верхняя завязь соединяется своим основанием с цветоложем (косточковые породы). Средняя завязь до половины срастается с цветоложем, а верхняя ее половина остается свободной (земляника). Нижняя завязь образуется в результате срастания оснований чашечки, венчика и тычинок (семечковые породы — яблоня, груша, айва, рябина, боярышник; ягодные культуры — смородина, крыжовник; орехоплодные — грецкий орех, лещина).
Поскольку строение вегетативных и цветковых почек различно, то и фенологические фазы первоначального развития у них неодинаковы. Вегетативная почка проходит следующие фенофазы: набухание, раздвигание почечных чешуй, появление зеленого конуса, образование листочков, появление розетки полноразвитых листьев, начало роста побега, усиленный рост побега, затухающий рост побега, закладка пазушных почек, сбрасывание листьев. Цветковая же, или смешанная, почка проходит такие фазы: набухание, начало роста, распускание почки, выдвижение соцветий, обособление бутонов, появление венчиков, расхождение лепестков, цветение, опадение лепестков, завязывание плодов, смыкание чашелистиков, рост плода, созревание плода.
Последовательность в распускании вегетативных и цветковых почек у различных пород неодинакова, например у миндаля, абрикоса, персика, лесного орешника (лещины) цветковые почки распускаются немного раньше вегетативных. У яблони и груши вначале распускаются цветковые почки, а немного позднее вегетативные.
Вегетативные почки развиваются в розетки листьев, или вегетативные побеги. Цветковые (смешанные) почки образуют цветки, плоды и замещающие побеги. Размеры листьев, возникающих из цветковых почек, зависят от запаса питательных веществ, отложившихся в прошлом году, питания растения в ранневесенний период, погодных условий и сортовых свойств. Образование цветков зависит от степени дифференциации почек, условий их перезимовки, питания дерева, температурного и воздушно-водного режима.
В процессе эволюции плодовые деревья выработали способность создавать большие резервы цветковых почек на случай сильных холодов, засухи и других неблагоприятных условий. На одном плодоносящем дереве яблони находится до 200 тысяч цветков. Из них только 5—10% дают полноценный урожай.
Продолжительность фенофазы распускания зависит, прежде всего, от температуры и воздушно-водного режима. Особенно большое значение имеет среднесуточная температура воздуха. Распускание и цветение обычно протекают при среднесуточной температуре 8—10°. Температура почвы не оказывает такого решающего влияния на цветение, как температура воздуха. Если, например, сравнить два дерева, у одного из которых почва приствольного круга покрыта мульчой, а второе свободно от мульчи, то, несмотря на то, что у первого дерева температура почвы ниже, цветут оба дерева одновременно.
Созревание почвы ранней весной имеет большое значение для своевременного начала роста активных корней, листьев и побегов. Если, например, в конце зимы приствольный круг дерева укрыть толстым слоем снега, то эта мера не ускорит цветения, а начало роста корней может задержать. Медленное таяние снега приводит к запоздалому развитию корней, а из-за этого надземные органы не разовьются вовремя, что затормозит развитие всего дерева.
Более подробные данные о морфогенезе почек и соцветий – Формирование почек и соцветий

В силу исторически сложившихся обстоятельств Талыш является обособленным и уникальным флористическим районом Кавказа, рефугиумом, где сохранились и произрастают многие редкие виды остаток доледниковой и более поздней флоры [1, 8].
Одним из этих растений является ольха почтисердцевидная. Изучая особенности строения почек ольхи сердцевидной, их морфогенез и другие вопросы, можно получить ценные сведения не только о биологии растения, но и о путях, которыми шла приспособительная эволюция исследуемого вида. Большую ценность представляют данные изучения морфогенеза почек у интродуцированных растений [2, 4, 5, 17].
Материалы и методы исследования
Материалом исследования – распространенный во флоре Азербайджана редкий и исчезающий вид Alnus subcordata S.A.Mey. Основная цель исследовательской работы заключалась в изучении цветения, плодоношения и семенной продуктивности ольхи почтисердцевидной. Изучение морфогенеза проводилось по методике Молчанова А.А., Смирнова В.В., Михалевской О.Б. [11], фенологические наблюдения по методике ГБС [13], ритм роста побега по методике Озолинчюс Р.В. [15], изучение биологических особенностей редких растений по методике Семенова Г.П. [16].
Результаты исследования и их обсуждение
Alnus subcordata C.A.Mey. (ольха почтисердцевидная) – однодомное растение. Мужские цветки ее собраны в повислые сережки, а женские соцветия представляют собой колоскообразные шишки, в пазухе плюсок которых развиваются женские органы [5, 17].
Исследования показали, что с момента заложения меристематического бугорка почки ольхи почтисердцевидной до развития плодущего побега, как и у многих древесных растений, проходит длительный период. Этот период можно условно разделить на три этапа.
Первый этап – заложение почки и формирование элементов – продолжается в течение 13 месяцев; второй этап – выдвижение почки в побег с листьями и формирование мужских и женских соцветий – длится 9-11 месяцев; третий этап – опыление, оплодотворение, развитие семени и плода – протекает в течение 10-11 месяцев.
В целом цикл развития почки от заложения до формирования в плодущий побег охватывает период до 3-х лет. Можно предположить, что такой растянутый на годы онтогенез почек у реликтовой ольхи является результатом адаптации вида к изменяющимся в сторону похолодания климатическим условиям.
Все эти факты указывают на былую вечнозеленность этого третичного вида, распространенного много лет тому назад, а ныне реликтового растения нашей флоры.
Наблюдения показали, что у ольхи почтисердцевидной почки двух типов – вегетативные и смешанные. Зимой и весной они выглядят совершенно одинаково, как морфологически, так и по внутреннему строению.
В феврале в нераспустившихся еще почках в пазухе почти всех зачаточных листьев, заключенных в этой почке, уже есть бугорки почек (т.е. почки в почках), которые состоят из меристемы и двух покровов. Весной по мере выдвижения побега из материнской почки и развития листьев увеличиваются в размерах и почки в пазухе листочков. За год эта пазушная почка превращается в материнскую и состоит из 5-7 листьев, каждый из которых защищен двумя прилистниками. В пазухе каждого такого листа почки снова появляется новый бугорок. Эти новые почки уже дочерние по отношению к тем, в пазухе которых они появились и внучатые по отношению к тем, внутри которых год тому назад появилась их материнская почка.
Прилистники наружного листа, которые по времени появления самые старые в почке, более грубые. Прилистники более внутренних листьев относительно нежные, покрыты клейким веществом, уберегающим, по-видимому, их от холода. Этой же цели служат и волоски, густо покрывающие уложенные, очень складчатые зачаточные листья.
Таким образом, почечные элементы у ольхи почтисердцевидной защищены с одной стороны крупными и рано опадающими, по мере весеннего прорастания побега, прилистниками (которых по 2, у каждого из них по 5-7 листочков), с другой стороны клейким веществом и опущением (рис. 1).
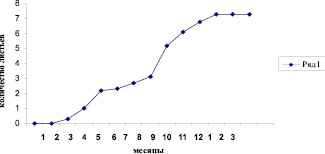
Рис. 1. Развитие листьев – из почки Alnus subcordata
В марте с наступлением теплых дней почка ольхи почтисердцевидной начинает распускаться. Готовый укороченный побег заключенный в почку с марта по июнь стремительно вытягивается (от 1 до 30 см) (рис. 2).
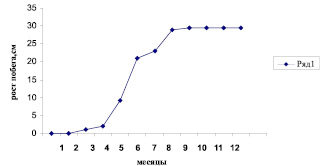
Рис. 2. Развитие побега Alnus subcordata из почки
Только в начале лета на отдельных побегах в пазухе последнего листа наблюдается формирование сначала мужских, а через 2-3 недели в верхушечной части этого же побега и женских соцветий (рис. 3).
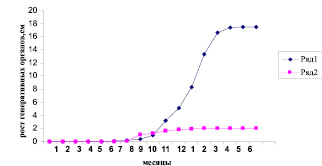
Рис. 3. Развитие мужских и женских соцветий Alnus subcordata
Одновременно нами проводилось изучение биологии цветения и учет плодов. По срокам начала и окончания цветения ольхи почтисердцевидной нами условно выделены на: а) ранее зацветающие и б) ранее отцветающие. Продолжительность цветения составила 20 дней; отметим, что похолодание ведет к удлинению периода продолжительности цветения исследуемого вида. Осыпание цветков является его биологической особенностью; так, у двуполой ольхи почтисердцевидной мужские сережки после высыпания из них пыльцы осыпаются как выполнившие свою биологическую функцию.
Наблюдения показали, что у рано зацветающих видов о.почтисердцевидной наблюдается и ранее созревание плодов. Не все появившиеся цветки сохраняются на побегах до полного формирования из них плодов, часть постепенно осыпается [14]. Процент образования плодов составил 87 %. В одной шишке исследуемого вида насчитывается 130-150 семян. Результаты исследования показали, что у 34-летний о.почтисердцевидной можно сосчитать около 17880 шишек и 2395920 семян. Общеизвестно, что образование полноценных семян у растения, в особенности у интродуцента, является важным показателем [7, 9, 12].
Знание сроков созревания плодов и семян имеет важное практическое значение при прогнозировании урожайности и организации непосредственного сбора и заготовки семян. Согласно нашим исследованиям установлено, что условия Апшерона вполне пригодны для их существования. Наблюдения показали, что для полного цветения, плодоношения и формирования жизнеспособных семян у исследуемых видов, путем реинтродукции и культивирования возможно сохранение, а также постепенное расширение ареала этого вида.
Нормальная репродуктивная ситуация ольхи свидетельствует о том, что причина узколокального местообитания не является какой-либо особенностью в биологии цветения и плодоношения.
Таким образом, проводимые нами исследования по изучению фенологии, особенностей внутрипочечного развития, ритмики и продолжительности отдельных фенофаз, цветения, плодоношения и семеновыведения о.почтисердцевидной дают основание считать интродукцию ее в условиях Апшерона успешной.
Формирование почек и соцветий – Закладка и развитие соцветий в зимующих почках винограда
Содержание материала
Значительно больше исследований посвящено закладке и формированию соцветий в почках винограда, причем они направлены в основном на выяснение времени заложения, роста и развития соцветий, влияния различных факторов и т.д.
Первые исследования дифференциации почек принадлежат Martelli (1892) и Miiller-Turgau (1892). Они показали, что зачатки первичных гроздей можно наблюдать уже в августе и что дифференциация соцветий начинается, когда почки находятся в зеленом состоянии. Исследования в этой области особенно усиливаются после тридцатых годов. Так, Snyder (1933) изучал дифференциацию почек сорта Конкорд, Barnard и Thomas (1933, 1937) — развитие соцветий в почках сорта Султанина в условиях Австралии, Winkler и Shamsettin (1937) — развитие этого же сорта в условиях Калифорнии. Обстоятельные исследования проводили также Breviglieri (1956), Khalil (1961) в Италии и др.
Сроки закладки соцветий в зимующих почках по длине побега и их плодоносность
Сроки закладки соцветий в зимующих почках винограда изучались многими авторами. По данным Winklern Shamsettin (1937), в Дэвисе (Калифорния) первые признаки дифференциации плодоносных почек в нижних частях побегов отмечаются в начале июня. До конца июня формирование соцветий отмечалось в почках 15-го глазка.
Формирование соцветий в Австралии и Южной Африке начинается с середины октября до начала декабря (Barnard, 1932; Barnardn Thom a s, 1933; Реrо1d, 1927). Во Франции дифференциация соцветий начинается примерно с середины июня и заканчивается в середине августа (Н и – g 1 i η, 1958), а в Италии — дней на 20—30 раньше — 16—18 мая (Breviglieri, 1956).
Исследования Титовой-Молчановой (1951, 1952) в условиях Средней Азии показали, что закладка зачатков соцветий в почках начинается почти одновременно у сортов Пино черный, Саперави, Ркацители, Ак кишмиш и Тайфи розовый — во второй половине мая и совпадает с периодом цветения винограда. Подобные результаты получил Дикань (1972).
К аналогичным выводам приходит также Менагарашвили (1957), изучавший вопрос о закладке соцветий в глазках сорта Ркацители в условиях Грузинской ССР. Науменко и З. В. Колесник (по сообщению Корнейчука и Плакиды, 1959) также установили, что зачатки первых соцветий изучаемого ими большого набора сортов (Анжевин Оберлен, Шасла золотистая, Каберне, Гаме черный, Сенсо, Серексия, Рислинг, Алиготе, Ркацители, Пино серый), за исключением Нимранг и Карабурну, образуются почти одновременно. Колесник и Колесник (1969) сообщают, что бугорки соцветий в районе Одессы наблюдались в конце мая — начале июня.
Титова – Молчанова (1951, 1952) установила, что у сортов, у которых рост побегов прекращается раньше (Пино черный, Ркацители и Саперави), закладка зачатков первых соцветий заканчивается в более сжатые сроки — примерно в течение месяца (в конце мая и в первой половине июня).
У сортов с более длинным вегетационным периодом (Тайфи розовый и Ак кишмиш) наблюдалось некоторое отставание закладки зачатков соцветий — наиболее интенсивно этот процесс проходил в конце июня. На этом основании автор приходит к выводу, что “периоды наиболее интенсивной закладки зачатков в почках у разных сортов не совпадают”.
Степень дифференциации и плодоносности почек по длине побега неодинакова. Более плодоносны и лучше дифференцированы соцветия в почках средней зоны побега (5—10-го узла), чем в нижней его части. Наблюдения З. Я. Титовой-Молчановой по установлению длины зародышевого побега и числа узлов на нем показали также неодинаковую степень дифференциации почек по длине побега. Глазки в средней зоне побега крупные, в них больше замещающих почек, а в главных почках этих глазков на зародышевом побеге больше узлов и лучше развитых зачатков соцветий.
Тютюник (1952), изучавший развитие зимующих почек винограда и закладку в них соцветий в средней полосе СССР, также пришел к выводу, что разница в сроках закладки и дифференциации соцветий в центральной почке (10-го глазка) у различных сортов довольно значительна и достигает 20—40 дней.
Из приведенных данных видно также, что от закладки бугорка до дифференциации первого соцветия проходит около месяца (23—33 дней). Период дифференциации второго соцветия несколько короче (19—30 дней). Закладка бугорка и дифференциация второго соцветия начинается примерно на 10— 12 дней позже начала закладки первого соцветия.
Исследования, проведенные в условиях Болгарии Стоевым и Никовым (1956), показали, что закладка бугорка начинается в последней декаде мая, а самих соцветий — в конце мая — начале июня. Разница в начале закладки соцветий ранних и поздних сортов составляет примерно 10—14 дней, причем период заложения соцветий всегда приурочивается к началу периода цветения сорта. Особо характерен в этом отношении сорт Димят. По сроку созревания ягод он принадлежит к почти поздним сортам, однако цветение у него начинается рано и соцветия закладываются также рано.
Таблица 3
Динамика закладки соцветии в центральных почках глазков, %
Зоны побега (узлы)
Следует отметить, что обнаруженная в начальные моменты разница в закладке соцветий у разных сортов через месяц почти исчезает (табл. 3).
С тем чтобы установить темпы закладки соцветий по периодам протекания этого процесса, авторы сопоставляли сформированные соцветия в начальные 20 и последние 40 дней. Соответствующие данные приведены в табл. 4.
Таблица 4
Процент соцветий, сформированных в интервале 9—30 июня, 1 июля — 11 августа в различных секторах побега (1—15-й узел)

* К 9 июня не было соцветий.
вается больше соцветий, чем позже (1 /VII—11/VIII) за более длительный срок. К аналогичным выводам приходит также Менагарашвили (1957), отмечая, что максимальное количество соцветий образуется в период цветения до первой половины июля. В период торошения закладывается много соцветий, но все же меньше по сравнению с первым периодом. Таким образом, закладку соцветий нельзя рассматривать как плавный процесс, протекающий равномерно в течение вегетационного периода. Наиболее интенсивно оно протекает в секторе 6, 10 и 15-го узлов, т.е. в средней и выше средней частях побега. Как видно из данных, в 60—65% зимующих почек 15-го узла сортов Димят, Памид и Болгар закладка соцветий произошла примерно за 10 дней.
Как отмечалось выше, по данным Титовой-Молчановой (1952а) формирование почек происходит тем быстрее, чем выше они находятся от основания побега. В соответствии с этим и дифференциация соцветий осуществляется быстрее и лучше, в результате чего высоко расположенные почки более плодоносны.
Период наиболее интенсивной закладки соцветий считается критическим в годичном цикле виноградной лозы, причем критический период роста и развития почек наступает разновременно для почек побега, передвигаясь последовательно с базальных к средним и выше средних зонам побега.
Процесс роста соцветий и полной их дифференциации продолжается и после периода интенсивного темпа их закладки. Исследования Стоева и Н и к о в а (1956) в этом направлении показали, что находящиеся у основания побега глазки вначале имеют более крупные соцветия. Несколько позже, однако, рост соцветий в расположенных выше почках значительно усиливается, благодаря чему уже к середине августа наиболее развитыми оказываются соцветия в секторе 10—15-го узла (табл. 5).
Более крупные соцветия в почках средней и выше средней зон побега установлены множеством авторов в различных виноградарских районах и у самых разнообразных сортов. Это можно считать одной из наиболее характерных закономерностей биологии виноградной лозы. Наблюдаемые позже во время вегетации различия в величине гроздей на побегах разных секторов прошлогоднего побега существуют заранее в глазках, еще во время закладки и дифференциации соцветий, и этим в значительной степени определяется характер плодоношения в следующем году.

Рис. 2. Коэффициент плодоношения (кп) по длине побега
1 — Димят; 2 — Памид; 3 — Болгар; 4 — Мавруд (а-м)
Таблица 5
Динамика развития соцветий* в почках винограда

* Величина соцветий в микронах.
Немного после формирования первых происходит закладка вторых и последующих соцветий в почках. По данным Стоева и Никова (1956) для этого требуется 20—25 дней, причем этот срок длиннее для низкорасположенных глазков и короче для средних и выше средних почек. Способность к формированию более одного соцветия в почках ярче выражена в расположенных выше глазках, поэтому коэффициент плодоносности более высокий (рис. 2). Лучшая дифференциация соцветий, по-видимому, объясняется лучшим питанием почек в результате повышенной ассимиляционной активности листьев средних частей основного и бокового побегов (Стоев с сотр., 1952; Стоев с сотр., 1961; Стоев с сотр., 1966; Колесник, 1953).
О закладке соцветий в замещающих почках сообщает также Плакида (1953). К этому выводу она пришла, изучая плодоносность замещающих почек после удаления весной всех главных почек глазка. Как отмечает автор, удаление центральных почек совершенно не сказалось на величине урожая и среднем весе грозди сорта Шасла, а в сильной степени отразилось на урожайности сорта Муската белого.
Соцветия закладываются также в зимующих почках на пасынках. Как показали исследования Стоева и Никова (1956), закладка соцветий в почках на пасынках начинается в первой декаде июля, причем раньше на пасынках нижних и средних ярусов основного побега (1—4-го и 7—9-го узла), а позже (в последней декаде июля) — и на пасынках, развитых выше средней зоны основного побега. По длине пасынка в нижней части побега (1—4-го узла) обнаружена акропетальная последовательность соцветий. На пасынках в секторе 7—9-го узла основного побега плодоносность почек не меняется метамерно, а на пасынках в секторе 12—15-го узла увеличивается базипетально. Авторами установлено также, что на пасынках процент плодоносных почек (почек с соцветиями) несколько ниже, чем на основном побеге. Это не совсем согласуется с наблюдаемым очень часто в производственных условиях более высоким коэффициентом плодоносности пасынков. Это несоответствие можно объяснить предположением, что микроскопическим путем не всегда точно учитываются соцветия в почках пасынков, которые, как правило, меньшей величины, чем в почках основного побега. Можно полагать также, что заложенные летом бугорки в почках пасынков подвергаются дальнейшему развитию и дифференциации в ранневесенний период или же, что проведенные нами наблюдения приостановлены слишком рано (1/IX). Плодоносность почек на пасынках установлена также Milosavlievic и Pemovski (1977) и другими авторами.
После искусственного проращивания зимующих почек в год их формирования на выросших побегах вновь образуются сложные почки (глазки), а в них закладываются вполне нормально развитые соцветия. Стоев и Ников (1956), удаляя все точки роста на побеге после 15 июня, наблюдали закладку соцветий на развившихся побегах из зимующих почек, и на них формировались плодоносные пасынки. Таким образом они установили возможность закладки соцветий у четырех генераций зимующих почек: на основном побеге, на развитых на нем пасынках, на основных побегах II генерации, выросших преждевременно из зимующих почек, и на сформированных на них пасынках II генерации.
Несколько позже Никову (1964) удалось получить в течение одного вегетационного периода четыре генерации плодоносных побегов из зимующих почек (последние три генерации — за счет преждевременно проросших зимующих почек). При этом в зимующих почках на побегах всех генераций закладывались соцветия, что видно из данных табл. 6.
Таким образом, способность виноградной лозы формировать почки на основных побегах и на пасынках и закладывать в них соцветия очень велика. Это имеет большое теоретическое и практическое значение. Факт появления четырех генераций плодоносных почек на основных побегах объясняется и тем, что этот процесс в последующих генерациях протекает значительно интенсивнее и в более сжатые сроки. Коваль и Матвеев (1972) считают, что любая почка, несмотря на ее расположение, способна при определенном сочетании условий превратиться в плодоносную.
В благоприятных условиях нормальные соцветия закладываются также в почках жирующих побегов, развившихся из спящих почек и побегов из замещающих почек глазков.
Многие исследователи раньше считали жирующие побеги неплодоносными. Например, Vialan Vermorel (1910) отмечают, что бесплодие жирующих побегов V. vinifera (за небольшими исключениями), V. labrusca, V. aestivalis и др. постоянно (неизменно). По мнению Вranas с сотр. (1946) и Вranas (1947), почки жирующих побегов также бесплодные.
Таблица 6
Закладка соцветий* у четырех последовательных генераций основных побегов

* Учет проводился в почках на 5-м узле побега.
Некоторые исследователи рассматривают почки жирующих побегов как бесплодные (Dumler, цит. по Нug1in, 1955), другие же (Wо1f, 1931; Weger, Moog, 1949; Zilling, Heymer, 1950) наблюдали плодоносные глазки у жирующих побегов сорта Рислинг. Sсhеu (1955) отмечает, что иногда можно наблюдать плодоносные глазки у жирующих побегов некоторых сортов, таких, как Рислинг и Мюллер-Тюргау. Бесплодными считали почки жирующих побегов также ряд болгарских авторов (Неделчев, Кондарев, 1967).
Наряду с этим в последние годы все больше утверждается взгляд, что почки жирующих побегов не менее плодоносны, чем все остальные почки на побегах, развитых из прошлогодней древесины. Например, Турянский (1950 б) приводит данные, из которых видно, что не имеется существенной разницы в плодоносности почек побегов, развившихся из главных и замещающих почек основного побега, выросшего из спящих почек. Гузун и Семин (1963) даже установили наиболее высокую плодоносность почек на побегах, выросших из спящих почек.
Существенный вклад в изучение вопроса о плодоносности почек на жирующих побегах внес Нug1in (1955). Он изучал среднее число соцветий в почках жирующих побегов в условиях полного удаления однолетних и двухлетних побегов (опыт в 1954 г.) и при совместном выращивании выросших из спящих почек и нормальных побегов, выросших из почек прошлогоднего побега (опыт в 1955 г.). Наблюдения показали, что разница в плодоносности почек, развивающихся на нормальных и жирующих побегах сортов Knipperld, Sylvaner, Muscat Ottonel, Chasselas, Riesling, Pinot gris, Pinot blanc, Gewurz- traminer, Auxerrois, незначительна. На основании этих результатов автор заключает, что почки жирующих побегов обладают такой же плодоносностью, как и нормальные побеги, и аналогичной силой роста. К подобным выводам пришли также Павлов (1963) и Georgescu и I1iе (1971).
Источники:
http://plodovyy-sad.ru/obshchie-zakonomernosti-razvitiya-privitykh-plodovykh-derevev/obrazovanie-i-razvitie-tsvetkovykh-pochek-u-kostochkovykh-kultur/
http://applied-research.ru/ru/article/view?id=9188
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/formirovanie-pochek-i-socvetiy-3.html




