
Пропускание солнечной радиации кроной винограда – Радиационный режим виноградника
Пропускание солнечной радиации кроной винограда – Радиационный режим виноградника
РАДИАЦИОННЫЙ РЕЖИМ виноградника
РАДИАЦИОННЫЙ РЕЖИМ виноградника, поступление и распределение энергии солнечной радиации в оптикобиологически системе, состоящей из расположенных определенным образом растений и частей растений с известными геометрическими и оптическими свойствами. В формировании Р. р. винограда решающую роль играет архитектура растений и структура виноградника. В шпалерно-рядовых виноградниках наибольшее количество радиации (Р) на единицу площади за день поступает на верхнюю сторону кроны. Кривая дневного хода Р для этой стороны симметрична относительно истинного полдня. Наиболее низкие интенсивности Р — на северной стороне шпалеры, куда поступает главным образом рассеянная Р с интенсивностями 0,1—0,2кал/см 2 .мин. На восточную и западную стороны за день приходится-примерно одинаковое количество Р. На восточной стороне максимум облученности наблюдается в 8—9 ч, на западной — в 16—17 ч. Пропускание Р внутрь кроны для боковых сторон наибольшее в утренние и послеполуденные часы, наименьшее — в околополуденные. Радиационное поле (РП) кроны в вертикальном слое характеризуется большой пестротой интенсивностей Р. На протяжении дня наиболее однородное РП с относительно низкими интенсивностями Р складывается в полуденные часы. В утренние и послеполуденные часы увеличиваются абсолютные значения Р и одновременно повышается пестрота РП в слое. По мере понижения слоя кроны количество отсчетов с низкими интенсивностями Р увеличивается.» Ослабление Р внутри кроны неодинаково для лучей различной длины волн. Наибольшее ослабление Р отмечается в области красных и синих лучей, наименьшее — в области зеленых и близких инфракрасных. Усредненные кривые вертикального распределения Р образуют на вертикальной шпалере 2 максимума — в 10 и 16 ч и один минимум — в околополуденные часы. На Т-образной шпалере дневной ход Р в кроне изменяется синхронно поступлению Р на горизонтальную поверхность. Поглощение Р виноградником в период вегетации характеризуется непрерывным увеличением среднедневных коэффициентов поглощения, которые достигают 0,7 для интегральной радиации, поступающей на весь виноградный массив. Увеличение ширины или высоты кроны ведет к пропорциональному (линейному) увеличению дневного поглощения Р. Специфика Р. р. виноградника проявляется в высокой облученности и аккумулирующей способности боковых сторон кроны в течение дня. Вследствие этого виноградники имеют более высокие коэффициенты поглощения Р, чем посевы при тех же размерах листовой площади на гектаре. Р. р. виноградника оказывает влияние на все стороны жизнедеятельности куста: фотосинтез, транспирацию, плодоношение, качественный состав биопродуктов в ягодах и др. Плодоносность почек повышается с увеличением дозы Р и не зависит от ее спектрального состава. При очень слабом доступе Р, а также при избыточной Р развитие ягод задерживается. При ограниченном доступе Р сильно подавляется рост губчатых, палисадных и эпидермальных клеток, пластинки листьев формируются более тонкие, чем у растений на полном свету.
Литература: Амирджанов А. Г. Солнечная радиация и продуктивность виноградника. — Л., 2000.
Радиационный режим виноградника – Пропускание солнечной радиации кроной винограда
Содержание материала
111.4. ПРОПУСКАНИЕ СОЛНЕЧНОЙ РАДИАЦИИ КРОНОЙ ВИНОГРАДА
Какова бы ни была величина поступающей солнечной радиации, распределение ее внутри кроны будет зависеть от характеристики пропускания солнечных лучей фитомассой. Коэффициент пропускания солнечной радиации ат определяется как отношение [361]:
aT = ql/Ql,
где qi — нисходящий поток суммарной радиации внутри кроны для г-того уровня; Q — приход радиации на горизонтальную поверхность.
Величина ат отдельного слоя вычисляется как среднее арифметическое: 
где aTi, aTt, ат—коэффициенты пропускания в отдельных
точках маршрута; и—число точек.
В табл. 10 в качестве примера приведены значения аГи для сорта Шабаш на вертикальной шпалере. Значения q„ в таблице не приводятся, так как они показаны на рис. 21.
Таблица 10
Коэффициенты пропускания ИР по ярусам кроны винограда на вертикальной шпалере (24 июля 1969 г.)
поступление ИР на
Верхний Средний Нижний
В утренние и вечерние часы коэффициенты пропускания довольно высокие. Однако надо принять во внимание, что в эти часы величина ат обусловливается главным образом радиацией, проникающей через боковые стороны кроны. По мере приближения к полуденным часам значения коэффициентов резко уменьшаются.
Коэффициенты пропускания, подобные представленным в табл. 10, позволяют получить радиационные характеристики дайной конкретной кроны, т. с. они имеют локальный характер. Более универсальной характеристикой ат является функция aT = f (LAI). В этом случае по значению LAI для данной культуры, независимо от места и времени проведения работы, может быть рассчитана величина ат или же решена обратная задача: найдена величина ILAI, которая с учетом поступления радиации и величины ат обеспечивала бы в том или ином участке кроны интенсивность радиации, достаточную для высокой фотосинтетической активности листьев.

Рис. 18. Зависимость функции пропускания суммарной ИР ат« от относительной площади листьев кроны винограда в течение дня. Сорта: 1 — Ркацители, 2 — Шабаш, 3 — Рислииг рейнский,
На рис. 18 показаны полученные нами экспериментально зависимости aTa = f (ILAIC) кроны куста в разные часы дня. Увеличению ILAIr соответствует понижение уровня пиранометра в кроне. При шести уровнях высоты кроны и восьми сериях для каждого срока получено 48 точек, характеризующих эту связь. В широком диапазоне дневных часов между аГи и ILAIr обнаруживаются тесные обратные линейные связи. Числовые характеристики уравнений приведены в табл. 11.
Таблица 11
Уравнения связи ajJILAIr и атф/ИА1г для кроны виноградного куста в разные часы дня (август) 
Представляло интерес сопоставить характер ослабления радиации фитомассой винограда при рядовой посадке с растительностью, имеющей сплошной покров. Из табл. 12 видно, что при LAl= величина ату кукурузы [366] более чем вдвое больше, чем у винограда при /JL4/r=l. При увеличении листового индекса до 2 разница сокращается, и при LAI = 3-г-7 значения am оказываются близкими. Относительно низкие аТа у винограда при малых ILAIr можно объяснить особенностями структуры верхнего слоя кроны, где часть побегов после окончания роста подвязывается к верхней проволоке горизонтально и в результате создается как бы эффект «экранирования».
Таблица 12
Изменениеaja в зависимости от величины LAI посева кукурузы [366] и ILAIr кроны винограда 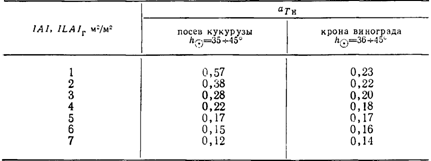
Как известно, в фитоценозах со сплошным покровом зависимость ослабления солнечной радиации от относительной площади листьев имеет вид экспоненты [306, 360]. Вместе с тем анализ экспериментальных данных показывает (рис. 19), что функцию aTn=f(LAI) можно удовлетворительно аппроксимировать при помощи двух прямолинейных отрезков: в интервале 1.Л1 = 0+2, где ослабление радиации наибольшее, и в интервале £Л/ = 2ч-7, где ослабление радиации происходит значительно слабее. Для винограда функции aTJILAIr и aT$/ILAIr для отдельных часов дня получены в интервале ILAlr = 2
l3, т. е. они приближаются ко второму участку экспоненты функции ати/LAl для ценозов сплошного покрова.
Аппроксимация криволинейных функций отрезками прямой имеет место и в других случаях. Отмечается, в частности, что предложенная Е. Рабиновичем (цит. по [360]) формула зависимости функции фотосинтеза от плотности потока падающей ФАР аппроксимирует световую кривую фотосинтеза также при помощи двух прямолинейных отрезков.
Как известно, радиационное поле внутри РП вследствие изменения соотношения между ФАР и БИКР 1 характеризуется неопределенностью состава суммарной ИР. Для перехода от коэффициентов пропускания ИР am к приближенным значениям коэффициентов пропускания ФАР атф Тоомингом [360] вычислены переходные коэффициенты (табл. 13). При /tc,>20° переход от аТи к атф не зависит от h.

Рис. 19. Зависимость функции пропускания суммарной IIP от относительной площади листьев в посевах кукурузы. а — пасмурно; б — ясно: Aq=40° (зачерненные точки). /i
=20° (светлые точки) (по [ЗОВ]); в — ясное небо, ЛГО —-10° (по [3(Ю]). Прерывистыми линиями показана аппроксимация кривой а^и ILAI двумя прямолинейными отрезками в диапазоне Ы/=0 + 2 и Ы/=2+7.
Расчеты показали, что зависимость пропускания ФАР от относительной площади листьев кроны винограда и времени дня представляется графически семейством параболических кривых, асимметричных по отношению к полдню (рис. 20). Эти кривые могут быть описаны математически.
1 БИКР — близкая инфракрасная радиация.

Рис. 20. Зависимость функции пропускания ФАР кроной винограда от времени дня т при разных значениях относительной площади листьев ILAIr. 
где ILAIr—относительная площадь листьев кроны; т — целые часы времени между восходом и заходом Солнца; At, 5, (i= = 0-:-5) — постоянные коэффициенты:
Л0= —6,73104; Л1 = 3,251871; Л2— —0,557435; Л3 = 0,04462833; Л4=—0,00171625; Л5 = 0,000026; В0=—1,528; ^ = 0,6513; Вг=—0,105375; В3=0,00821875; В4 =—0,0003125; Въ=0,0000046875.
Как известно, для характеристики радиационных потоков внутри посевов со сплошным покровом в зависимости от площади листьев (или индекса LAI) предложены различные математические модели, отражающие закономерности ослабления солнечной радиации внутри РП [100, 247, 365, 411, 471, 478,506].
Отмечая сложность теоретических формул и их приближенность, Росс и Тооминг [306, 360] предложили для описания режима солнечной радиации в посевах ряд полуэмпирических формул, в которых теоретические функции заменены эмпирическими постоянными.
Для количественного описания зависимости пропускания ФАР от ILAIr кроны винограда по экспериментальным значениям ити для отдельных часов дня с помощью переходных коэффициентов (табл. 13) были вычислены приближенные значения аТф для соответствующих значений lLAIr и часов дня т и по ним рассчитаны регрессионные связи атъ/ILAIr (см. табл. 11).
Последующая обработка данных, выполненная с помощью ЭВМ Мир-1, показала, что функция аТф = 1
Расписывая формулу в виде
аТф(1Шг т) = А0 + А±т + А2т2 + Л3т3 + Л4т4 + Аъ%ъ—
– -(Ва + В,т + В2т2 + Взт3 + В4т* + В5т5) /1Л/Г (III.6)
и подставляя в полученное выражение значения коэффициентов, получаем формулу для вычисления атф в зависимости от ILAIr и т.
Например, при ILAIr = 2 и т=10 получаем: аГф (ILAh = 2, х= 10) = —6,73104 + 32,51871—55,7435 + 44,62833 + 2,6— — (—1,528+6,513—10,5375 + 8,21875—3,125 + 0,46875) Х2 = = 79,74704—79,63704— (15,2005— 15,1905) X 2 = 0,11 —0,02 = 0,09.
Это же значение коэффициента получаем в случае использования соответствующего уравнения из табл. 11:
аГф = 0,11—0,01 X 2-0,11—0,02 = 0,09.
Применение формулы ограничивается условием, когда ширина кроны куста составляет 40—60 см, a /Li4/B = 3-r-5.
Амирджанов А.Г. Солнечная радиация и продуктивность виноградника – скачать книгу полностью

Скачать бесплатно книгу Амирджанов А.Г. Солнечная радиация и продуктивность виноградника
Читать онлайн книгу в форматах fb2, epub, pdf, mobi, lrf
Амирджанов А.Г. Солнечная радиация и продуктивность виноградника. –Л.: Гидрометеоиздат, 1980. –208 с.
Рост побегов винограда начинается после распускания почек и в зависимости от сорта и условий культуры продолжается до начала вызревания побегов или даже до наступления полной зрелости ягод. Наиболее интенсивно побеги обычно растут в период от распускания почек до начала цветения. К концу этого периода рост их достигает 60% и больше полной длины, а суточный прирост – 8–10 см. Снижение скорости роста побегов в период цветения не является обязательным. При некоторых условиях скорость роста возрастает во время цветения или же после его окончания и достигает 5–10 см в день [245, 483].
Междоузлие над листом растет в длину в течение 18– 20 дней и достигает максимального прироста, когда становится четвертым – седьмым; рост приостанавливается у восьмого–десятого междоузлия, считая от верхушки побега. В период цветения из сформировавшихся 19 листьев заканчивают свой рост 7–8 от основания побега, а 12 листьев достигают необходимой величины, чтобы отдавать ассимиляты другим частям растения [355].
У значительного числа сортов к моменту цветения побеги имеют по 14–16 узлов с листьями [170]. Удлинение побега происходит в основном путем вытягивания образовавшихся молодых клеток междоузлий (интеркалярный рост) и только отчасти за счет деления клеток верхушечной меристемы. На каждом побеге зона наиболее интенсивного вытягивания охватывает второе, третье и четвертое междоузлия, считая от верхушки. Рост нижележащих междоузлий постепенно замедляется и примерно на шестом-седьмом междоузлии совершенно прекращается. Одновременно с удлинением побега происходит его утолщение: в верхней зоне только за счет увеличения размеров клеток, а ниже – в результате возникновения вторичной меристемы (камбия), формирующей новые концентрические слои флоэмы и ксилемы. Одновременно с ростом побегов увеличивается количество листьев на побеге, их размер, изменяется облиственность побега и всего куста.
Стимулирующее действие света на накопление красящих веществ в ягодах винограда отмечает Бегунова. У сортов Каберне Совиньон и Саперави в ягодах гроздей, лишенных доступа света, появление окраски задерживалось на 8 суток, а количество красящих веществ в ягодах, созревших в темноте, было почти в два раза меньше, чем в ягодах на свету.
Обстоятельная работа по выявлению роли света в окрашивании ягод винограда выполнена Уивером и Мак Кьюном. Грозди сортов Токай, Султанина розовая и Эмперор в момент завязывания ягод изолировались от света и вплоть до уборки урожая находилась вне действия прямых солнечных лучей. Затенение практически не повлияло на созревание: среднее содержание РСВ и кислот в ягодах было соответственно 17,5 и 0,90% у растений на полном свету и 17,7 и 0,93% –У затененных. У сорта Токай при полном отсутствии света ягоды не окрасились. При частичном затенении грозди окрашивалась та ее часть, которая подвергалась действию солнечных лучей.
В другом опыте у сорока сортов винограда ягоды изолировались от света до начала их окрашивания. В момент уборки урожая у всех сортов, за исключением сортов Токай и Султанина розовая, ягоды затененных гроздей имели такую же окраску, как и на полном свету. Затенение не оказало влияния на содержание РСВ и кислотность ягод. По мнению авторов, действие света на окрашивание винограда зависит от пигментного состава ягод. В частности, для образования пигментов сорта Токай с Rf 0,23 и 0,14 свет необходим. Группа пигментов с Rf 0,28 и 0,10 может развиваться в темноте. Для большинства сортов действие света в период от завязывания ягод до начала окрашивания не было обязательным.
В исследованиях Наито [475, 476] многие сорта винограда со слабой окраской ягод обнаружили высокую требовательность к свету. В то же время группа сортов с темноокрашенной ягодой не имела различий по степени окраски ягод, если даже грозди созревали в темноте.
По данным Кливера [440], реакция сортов Кардинал и Пино гри на интенсивность света неодинакова. У сорта Кардинал при температуре 20 °С и освещенности 5–12 тыс. лк ягоды окрашивались также или несколько интенсивнее, чем при той же температуре и освещенности 25–50 тыс. лк. У сорта Пино гри независимо от температуры воздуха ягоды окрашивались всегда слабее при низкой освещенности.
Таким образом, между освещенностью и качественным составом биопродуктов ягод обнаруживаются довольно сложные зависимости. В одном случае действие света на содержание биокомпонентов ягод проявляется отчетливо (содержание яблочной кислоты), в другом – менее ясно (содержание сахаров и РВС). Неоднозначно проявляется действие света и на содержи ни и красящих веществ в ягодах. Сложности эти определяются, очевидно, тем, что хотя в экспериментах изучается действие света, его трудно вычленить из совокупного действия других факторов. А их может быть много: географическое местоположение виноградника, определяющее поступление суммарной солнечной радиации, сортовые особенности (особенно проявившиеся в окрашивании ягод), густота кроны, соотношение между листовой поверхностью и урожаем и др. Немаловажное значение имеет и то обстоятельство, что в опытах, проводимых в естественных условиях, обычно регулируется освещенность, но сохраняются естественные условия температурного режима воздуха, роль которого в формировании качества ягод показана всей многовековой практикой культуры винограда. Все это говорит о том, что для практических целей необходимо осуществлять работы для конкретизации действия светового фактора на качественный состав биопродуктов ягод в данной конкретной географической зоне на существующем сортовом материале, учитывая при этом, что качество ягод формируется и результате совокупного действия многих факторов, как внешних, так и обусловленных состоянием растения.
Не получается скачать и прочитать книги в форматах djvu, pdf, rar? Прочтите подсказки
Источники:
http://sortov.net/info/radiacionnyy-rezhim-vinogradnika.html
http://vinograd.info/info/solnechnaya-radiaciya-i-produktivnost-vinogradnika/radiacionnyy-rezhim-vinogradnika-5.html
http://kachat-knigi.ru/knigi-vinograd/2010-05-08-Solnechnaya-radiatsiya-produktivnost-vinogradnika.htm




