
Роль органических кислот при технологии и выдержке вина – Метаболизм органических кислот дрожжами при брожении
Роль органических кислот при технологии и выдержке вина – Метаболизм органических кислот дрожжами при брожении
Роль органических кислот при технологии и выдержке вина – Метаболизм органических кислот дрожжами при брожении




Обмен веществ (метаболизм) дрожжей
Микроорганизмы осуществляют свои жизненные функции, главным проявлением которых является рост и размножение за счет многочисленных непрерывно происходящих и взаимосвязанных химических реакций, обычно называемых обменом веществ или метаболизмом. Метаболизм представляет собой два типа процессов, а именно:
анаболизм или ассимиляция характеризуется синтетическими реакциями, при которых потребляется энергия, организмы принимают из среды вещества (питание), эти вещества они способны переработать в продукты, компенсирующие потери, вызванные потреблением и расщеплением, и пригодные для образования новых веществ для роста и размножения;
катаболизм или диссимиляция, т. е. сумма реакций разложения, богатых энергией высокомолекулярных веществ, на вещества простейшие, бедные энергией, а освобождающаяся энергия потребляется на процессе ассимиляции.
Метаболизм сахаридов
В обмене веществ дрожжей и всех живых систем вообще большое значение имеют сахара (сахариды), при расщеплении которых получается энергия для физиологических процессов и для биологического синтеза многих веществ. Расщепление сахаров (гликолиз) является одним из основных процессов. Если он протекает при прямом участии кислорода воздуха, т. е. при аэробных условиях, таких, как окислительный обмен веществ, речь идет о процессе, обычно называемом дыханием (respirace). При анаэробных условиях брожение (ферментация) протекает как антиокислительный обмен веществ.
Дыхание и брожение – это ферментативные процессы, сложный механизм которых в начальных стадиях подобен. Они протекают вначале через ряд промежуточных стадий, а затем на определенной стадии разделяются и идут к конечным продуктам своими собственными путями. При дыхании конечным продуктом являются углекислый газ и вода
и при брожении спирт (этанол) и углекислый газ
Количество энергии, освободившейся при этих катаболических (диссимиляционных) процессах, зависит от степени окисления. Чем выше теплота сгорания образующихся продуктов, тем меньше энергии освободилось для клетки. Метаболизм сахаридов (углеводов) и его отношение ко всему обмену веществ выражает следующая схема [5]:
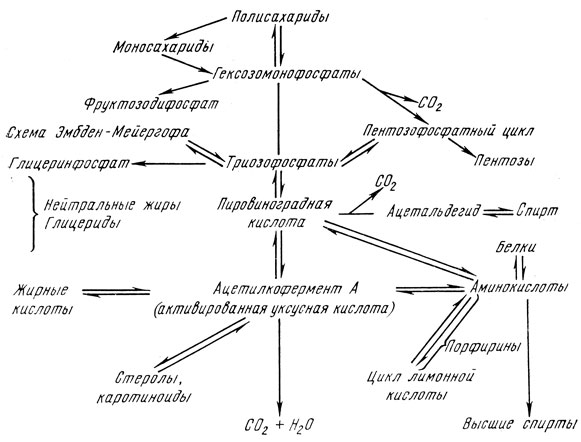
Полисахариды и олигосахариды (мальтоза, сахароза) или сначала расщепляются на моносахариды и фосфорилируются, прямо переходят в фосфаты гексозы. Последний из них фруктозо-1,6-дифосфат расщепляется на два фосфорилированных С3-сахара (по схеме Эмбден-Мейергофа) или уже на предшествующей стадии глюкозо-6-фосфат дегидрогенизируется на 6-фосфоглюконовую кислоту и путем окислительной декарбоксилации перейдет в пентозофосфат, который является голозидным компонентом рибонуклеиновых кислот. Фосфаты пентозы в так называемом пентозофосфатном цикле переходят через гептозы, гексозы, тетрозы в фосфаты триоз и гексоз, через которые присоединяются к схеме Эмбден-Мейергофа.
Из триозофосфата (дигидроксиацетонфосфата) образуется в качестве побочного продукта глицерин, основное вещество для синтеза нейтральных жиров и фосфатидов.
Основной путь расщепления сахаридов ведет от триозофосфата через разные промежуточные стадии к пировиноградной кислоте. При декарбоксилации и восстановлении образуются основные продукты анаэробного гликолиза, т. е. этанол и углекислый газ. Кроме того, пировиноградная кислота, отдает свой углеродный скелет аминокислоте аланина, который образуется в результате трансаминирования.
При аэробных условиях протекает также процесс окисления, в результате которого образуется “активированная уксусная кислота”. При скоплении ацетиловых остатков активированной уксусной кислоты образуются жирные кислоты, стеролы, каротиноиды и одновременно образуется углеродный скелет ряда важных аминокислот. Эти процессы обратимы и поэтому ацетилкофермент А является важным промежуточным веществом расщепления, перестройки и построения сахаридов, белков, жиров, стеролов, каротиноидов и других веществ.
Окончательное превращение ацетиленового остатка в углекислый газ и воду осуществляется в цикле лимонной кислоты, благодаря чему клетка приобретает основную часть энергии. Из промежуточных продуктов α-кетоглютаровой и щавелевоуксусной кислот при аминировании и трансаминировании образуются глютаминовая и аспарагиновая кислоты.
При пивоваренном брожении основным и преобладающим процессом является анаэробный гликолиз (брожение) по схеме Эмбден-Мейергофа-Парнасова, при котором в качестве основных продуктов образуются этанол и углекислый газ. В присутствии кислорода на процесс брожения накладываются аэробные процессы (дыхание), в частности в стадии пировиноградной кислоты цикл лимонной кислоты (Кребса) или в стадии глюкозо-6-фосфата-пентозофосфатный цикл.
При низовом пивоваренном брожении приблизительно 98% сахара сбраживается и 2% используются на процесс дыхания.
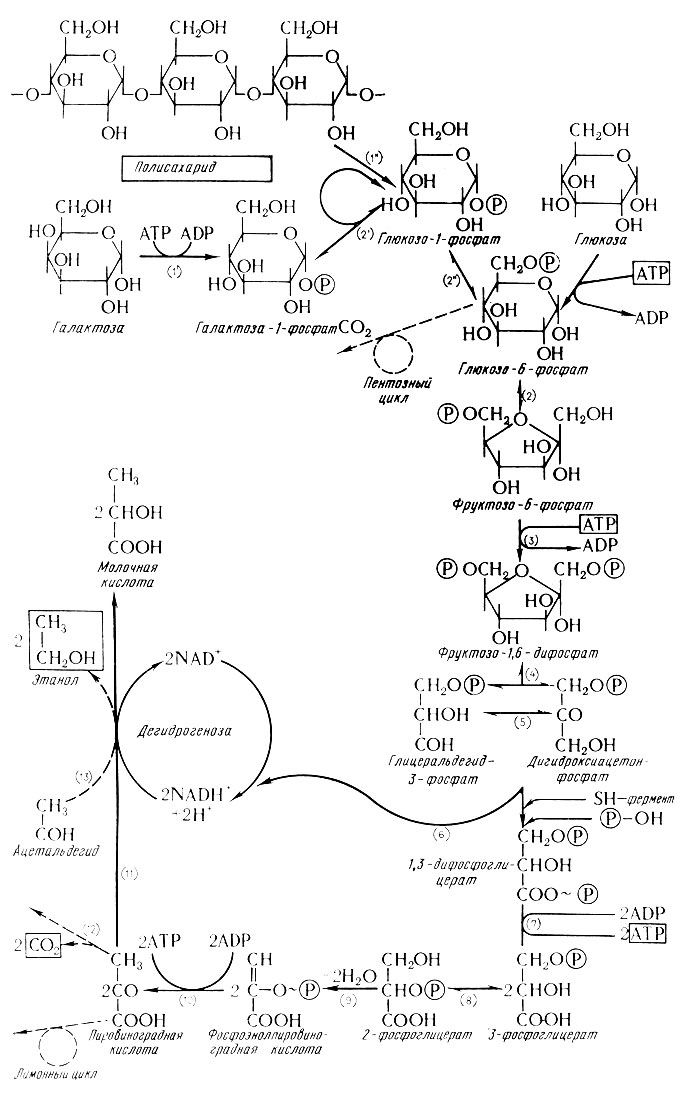
Спиртовое брожение (схема Эмбден-Ме йергофа)
Спиртовое брожение очень сложный процесс, при изучении которого постепенно выяснилось, что он протекает как цепь ре-акций, связанных фосфорилирующим механизмом.
В первой фазе брожения (стадия фосфорилирования) образуются разные фосфаты гексоз. Последний из них (фруктозо-1,6- дифосфат) распадается на два триозофосфата: дигидроксиацетонфосфат и глицеральдегид-3-фосфат. Глицеральдегид-3-фосфат дегидрогенизируется при вмешательстве никотинамидадениндинуклеотида (НАД) и одновременно фосфорилируется в 1,3-фосфорглицериновую кислоту, которая последующей дегидрогенизацией и дефосфорилизацией переходит в пировиноградную кислоту. Кислота в свою очередь декарбоксилируется в углекислый газ и ацетальдегид, который в последней фазе восстанавливается в спирт (этанол). Весь ход спиртового брожения виден по при-веденной по Де Клерку (6) схеме, которая составлена на основе имеющихся данных, относящихся к изучению этого сложного процесса.
Сбраживаемость сахаров различна и зависит от вида сахара и от свойств дрожжей. Сначала считали, что прямо сбраживаются только так называемые зимогексозы, т. е. глюкоза, фруктоза, маноза и условно (после приспособления) также галактоза, при этом все только в оптической D-форме. Олигосахариды считались сбраживаемыми только после расщепления на гексозы соответствующими ферментами. Например, низовые пивоваренные дрожжи содержат инвертазу (сахарозу, β-фруктозидазу-β-D-фруктофуранозид фруктогидролазу, ЕК 3.2.1.26), катализирующую расщепление сахарозы на глюкозу и фруктозу, далее мальтозу (α-глюкозидазу-α-D-глюкозид глюкогидролазу, ЕК 3.2.1.21), которая катализирует расщепление мальтозы на две молекулы глюкозы и, наконец, мелибиазу (α-галактозидазу-α-D-галактозид галактогидролазу, ЕК 3. 2. 1. 23), которая при одновременном присутствии сахарозы расщепляет трисахарид на все три его составляющие, т. е. галактозу, глюкозу и фруктозу. У верховых дрожжей мелибиаза отсутствует и поэтому они расщепляют рафинозу на мелибиозу и фруктозу, так что сбраживают только ее фруктозный компонент ( 1 /3).
В последнее время с помощью хроматографического метода подтвердилось существование трансфераз, способных переносить гексозные единицы с одной молекулы дисахарида или трисахарида на другую, или также внутри молекулы с одной связи на другую. Так, под действием трансглюкозидазы может образоваться например, из сахарозы трисахарид фруктозо-фруктозо-глюкоза или фруктозо-глюкозо-фруктоза. Образовавшиеся таким образом сахара под действием трансфераз расщепляются (сбраживаются) непосредственно ферментами брожения. Сбраживаемость сахаров зависит также от проницаемости клеточной оболочки дрожжей. Известно, что внутрь дрожжей проникают дитрисахариды, а высшие сахара и декстрины никогда.
Согласно суммарному уравнению брожения из 100 г глюкозы теоретически получается 51,1 г спирта (этанола) и 48,9 г углекислого газа. Практически выход всегда ниже и соответствует в среднем 48,6% этанола и 46,4% углекислого газа. Около 2% глюкозы дрожжи ассимилируют и остаток приблизительно 3% переходит в побочные продукты, т. е. ацетилдегид, глицерин, уксусную и молочную кислоту.
Цикл лимонной кислоты
В начале брожения сусло всегда содержит определенное количество растворенного кислорода воздуха, в присутствии которого дрожжи “дышат”. Тем самым изменяется первоначальная схема анаэробного метаболизма. Восстановление ацетилдегида замедляется, так что начинается скапливаться пировиноградная кислота, незначительный излишек которой аэробно декарбонизируется в цикле лимонной кислоты, или в трикарбоксиловом цикле, называемом также по имени его открывателя циклом Кребса.
В реакцию включается следующий комплекс ферментов, катализирующий в присутствии кофермента А образование ацетил- кофермента А из очень небольшого количества пировиноградной кислоты. При его действии на щавелевоуксусную кислоту образуется лимонная кислота, а из нее постепенно кислоты цисаконитовая, L-изолимонная, щавелевоянтарная, α-кетоглутаровая, фумаровая, L-яблочная и, наконец, щавелевоуксусная, которая остается в цикле. Ее реакцией с пировиноградной кислотой начинается новый цикл.
Уже раньше было известно, что при полном исключении дыхания снижается жизнеспособность пивных дрожжей, а тем самым снижаются и их физиологические функции и бродильная активность. Вайнфуртнер [3] приводит, как вывод из новых исследований [15], сведения о том, что энергия, получаемая при дыхании, физиологически неравноценна энергии, полученной при брожении, поскольку без дыхания дрожжи постепенно утрачивают способность к размножению.
Пентозофосфатный цикл
В последнее время обнаружилось, что при дыхании дрожжей до 30% глюкозы расщепляется прямым окислением [16] в следующем так называемом пентозофосфатном или гексозомонофосфатном цикле по схеме Варбурга, Дикенса и Хорекера. При спиртовом брожении он накладывается на схему Эмбден-Майергофа в стадии глюкозо-6-фосфата, который окисляется в 6-фосфоглюконовую кислоту (через δ-лактон). При декарбоксилации через 6-фосфо-3-кетоглутаровую кислоту образуется рибулозо-5- фосфат. Последняя образует с рибозо-5-фосфатом седогептулозо-7-фосфат и глицеральдегид-3-фосфат, который возвращается в нормальную схему Эмбден-Мейергофа. Седогептулозо-7-фосфат расщепляется в эритрозо-4-фосфат и дигидроксиацетон. В присутствии глицеральдегид-3-фосфата образуется фруктозо-6-фосфат, который начинает следующий цикл.
Одним кольцом пентозофосфатного цикла одна молекула глюкозо-6-фосфата декарбонизуется в рибулозо-5-фосфат. При этом восстанавливаются две молекулы трифосфопиридиннуклеотида (ТПН), который снова должен окисляться кислородом воздуха.
Эффект Пастера
Основные изменения в метаболизме дрожжей, связанном с подачей воздуха в процессе брожения (анаэробный и аэробный процесс), первым заметил Пастер. Он обнаружил, что кислород подавляет брожение (образование спирта), однако ускоряет рост (размножение) дрожжей. Это явление известно как эффект Пастера. При аэробных условиях дрожжи не сбраживают (кроме пивных) сахар в этанол, а образуется только углекислый газ; при этом по сравнению с аэробным гликолизом потребление глюкозы намного меньше. По теории Линена [17], диапазон расщепления сахара регулирует концентрация неорганического фосфата и аденозиндифосфата (АДФ).
У пивоваренных культурных дрожжей эффект Пастера не наблюдается [18]. С технологической точки зрения наличие эффекта Пастера свидетельствовало бы о нарушении функций пивоваренных дрожжей.
Метаболизм азотистых веществ
Азотистые вещества в дрожжевых клетках постоянно обновляются. Одновременно с расщеплением протекает синтез белков. В качестве азотистых продуктов питания дрожжевым клеткам служат соли аммония, аминокислоты и низшие пептиды, если они проникают через оболочку клетки.
Для роста, обновления и восстановления живых систем особое значение имеют аминокислоты. Они образуются при введении аминогруппы в α-кетокислоты, которые являются промежуточными продуктами при расщеплении сахаридов. Преимущественную часть потребления аминокислот (около 75%) дрожжи осуществляют прямой ассимиляцией и почти весь остаток дезаминированием по схеме Эрлиха с образованием высших спиртов [19]. Для синтеза белков наиболее важна глютаминовая и аспарагиновая кислоты, аланин и амиды первых двух, т. е. глютамин и аспарагин.
Кроме аминокислот дрожжи ассимилируют также пептиды [20]. Из азота, ассимилированного из сусла дрожжами, около 11 % приходится на пептиды. Поэтому значение пептидов по сравнению с соответствующими аминокислотами меньше, а именно потому, что под действием экзопептидаз они сначала должны расщепляться в простые компоненты.
Внутри дрожжевых клеток аминокислоты подвергаются трансаминированию катализованному трансаминозами, переносящими аминогруппы с одной молекулы на другую. Так, например, аланин образуется путем реакции глютаминовой кислоты с пировиноградной. Высокомолекулярные азотистые вещества образуются конденсацией аминокислот при участии аденозинтрифосфата (АТФ). В пивном сусле аминокислоты являются более важным источником азота, чем аммониевые соединения [21]. Легко ассимилируемыми являются аргинин, лизин, метионин, глютаминовая и аспарагиновая кислоты. И, наоборот, пролин, валин и другие выделяются в среду.
Сусло всегда содержит достаточное количество азотистых веществ. Из 600 мг ассимилированного азота, содержащегося в 1 л сусла (12%-ное), при брожении дрожжи потребляют около 200 мг [22]. В стадии роста содержание азота в дрожжах быстро возрастает, пока не достигнет почти двукратной величины [23] и потом снова постепенно падает вплоть до исходной величины. Дальнейшее уменьшение содержания азота связано с тем, что дрожжи выделяют в среду от 5 до 33% из собственного объема азота, вероятнее всего в виде аминокислот и пептидов.
Метаболизм минеральных веществ
Общее значение минеральных веществ в клеточном организме вытекает из следующих функций:
они поставляют основные (неорганические) элементы для построения органических соединений (углерод, азот, сера и фосфор);
регулируют и поддерживают в организме определенные величины некоторых физических или физико-химических свойств, например прочность клеточной оболочки, осмотическое давление протоплазмы, ионное равновесие, pH, проницаемость клеточной оболочки, растворимость белков, активность ферментов и т. д.;
являются составной частью некоторых биологических катализаторов, главным образом ферментов (железо, медь), витаминов (кобальт), красителей (магний), активируют или ингибируют многие ферментативные реакции (магний, железо, медь, кальций, калий, натрий, кобальт, цинк и т. д.) и тем самым прямо вмешиваются в метаболические процессы.
Процессы, происходящие при формировании вин
В вине, как и в любой сложной биохимической системе, непрерывно происходят сложные физико-химические и биохимические процессы, позволяющие вину поддерживать свои жизненные процессы. Жизнь вина проходит через пять этапов: образование молодого вина, его формирование, созревание, старение и отмирание.
Образование молодого вина — это период от момента дробления винограда до завершения процесса брожения. Для первой стадии этого этапа, включающей в себя дробление, прессование мезги и осветление, характерны процессы экстракции и диффузии, а также первичные и вторичные окислительно-восстановительные процессы, происходящие под действием ферментов, активизирующиеся после дробления ягод и попадания их в сусло.
Вторая стадия образования молодого вина включает в себя сам процесс брожения сусла или мезги, в результате чего принципиально изменяются исходные свойства продукта (сусла) и в значительной мере — его состав. Наибольшим количественным изменениям подвергаются сбраживаемые сахара, которые при производстве сухих натуральных вин полностью используются дрожжами. В процессе спиртового брожения, помимо главных продуктов — этилового спирта и углекислоты, образуются вторичные продукты, играющие большую роль в создании вкуса и аромата вин. К ним относятся глицерин, янтарная и уксусная кислоты, ацетальдегид, 2,3-бутиленгликоль, ацетоин, лимонная и пировиноградная кислоты, изоамиловый и изопропиловый спирты, эфиры. К концу брожения молодое вино обогащается полипептидами и аминокислотами — продуктами автолиза дрожжей. Фенольные соединения, реагируя с белками вина, образуют таннаты, выпадающие в осадок. Кислоты сусла, так же как и вновь образующиеся при брожении кислоты, частично окисляются или подвергаются превращениям по циклу ди- и трикарбоновых кислот. Витамины в первый период брожения потребляются дрожжами, а после автолиза происходит обратный переход их в вино. Окислительно-восстановительный потенциал (ОВ) по мере сбраживания сусла уменьшается (с 355—485 мВ до 135), достигая минимума в период бурного брожения, когда происходит полное потребление кислорода.
Формирование вина включает период от окончания брожения до момента первой переливки, т. е. до отделения молодого вина от осадка дрожжей.
Установлено, что на качество вина существенно влияют условия и сроки выдержки вина над осадком. При свободном доступе кислорода аминокислоты дрожжей могут стать источником образования нежелательных продуктов— альдегидов и аммиака, придающих винам неприятные тона окисления. Настаивание вина на осадочных винных дрожжах (1—2% массы вина) при 0—10°С устраняет указанный дефект, так как в этих условиях дрожжи выделяют в вино ферменты, но не обогащают его растворимыми азотистыми веществами.
Наиболее важным для этапа формирования является процесс так называемого “биологического понижения кислотности вин”, сущность которого заключается в превращении яблочной кислоты в молочную под действием молочнокислых бактерий Lactobacillus Leuconostoc или Pediococcus. Так как молочная кислота по сравнению с яблочной менее диссоциирована и имеет более мягкий вкус, рН вина повышается, исчезает его “зеленая”, резкая кислотность, вкус смягчается.
Этапы созревания и старения вина включают время выдержки его в бочках (резервуарах) и бутылках.
При созревании, протекающем от 2 до 4 лет при доступе кислорода воздуха, вино приобретает стабильность и присущие типу органолептические свойства, т. е. становится “розливозрелым”. Старение вина происходит без доступа воздуха в течение 4—12 лет и более. В этот период полностью развиваются тонкий вкус и специфический аромат, свойственные старым винам определенного типа.
Изменения в составе и качестве вин при созревании обусловлены сложными физико-химическими превращениями, из которых наибольшее значение имеют процессы окислительно-восстановительного характера и сахаро-амин-ные реакции. Поэтому очень важно обеспечить необходимый доступ кислорода в процессе выдержки вина. Взаимодействие составных частей вина с кислородом способствует уменьшению его терпкости в результате окисления полифенолов.
Переводя в осадок легкоокисляемые соединения, кислород повышает тем самым стабильность вин и способствует формированию их органолептических свойств. Избыток поглощенного кислорода приводит к переокислению вина в результате окислительного дезаминирования аминокислот с образованием из них аммиака и альдегидов, придающих вину горький привкус. При значительном накоплении этих продуктов “переокисленный” тон вина переходит в “мышиный”. Причины окисленного тона вина окончательно не установлены. Некоторые исследователи объясняют появление этого дефекта присутствием диацетила.
Старение вина начинается с момента, когда окислительные процессы не могут больше улучшать вкусовые и ароматические свойства вина. Вкус и аромат старого вина создаются при низком ОВ-потенциале. Наиболее тонкий букет в выдержанном вине бывает при величине ОВ-потенциала не выше 200—250 мВ.
Считается, что наибольшую роль в улучшении органолептических свойств вин при старении играют реакции этерификации, при которых из спирта и кислот образуются сложные эфиры. Однако прямой зависимости между качеством вина и содержанием в нем эфиров не установлено. Более того, отмечено, что при избыточном накоплении уксусноэтилового эфира в вине наблюдается неприятный привкус (“штих”). При взаимодействии альдегидов и спиртов образуются ацетали, являющиеся ценными компонентами букета вин. На формирование букета благоприятно влияют аминокислоты и продукты их взаимодействия с сахарами — меланоидины. Реакции меланоидинообразования наиболее интенсивно протекают в специальных винах. Продукты распада Сахаров придают вину карамельные (малажные) тона. При их чрезмерном развитии в таких винах, как херес, мадера и мускаты, теряется свойственный типу вина вкус и аромат.
В результате реакций этерификации и конденсации на внутренних стенках посуды, в которой выдерживается вино, появляются осадки (рубашка), состоящие из полимеризованных красящих веществ и других соединений. Этот процесс особенно характерен для красных вин.
Отмирание вина является конечной стадией его развития. Оно связано с необратимым распадом основных его компонентов и всей системы в целом. Объективными признаками начавшегося распада вин являются выпадение красящих веществ в осадок и приобретение вином неопределенного цвета с грязноватым оттенком, потеря аромата и появление неприятного запаха разложения, резкое изменение и ухудшение вкуса, обусловленное разрушением спирта и органических кислот и накоплением продуктов распада.
Срок жизни вина в основном зависит от его типа, качества и условий хранения, но рано или поздно вино разрушается. Лучшие натуральные вина можно выдерживать до 30—35 лет, а наиболее экстрактивные высококачественные специальные крепкие и десертные вина — до 100 лет и более.
Изменение состава сусла в процессе брожения
Спиртовое брожение виноградного сусла связано с превращениями не только усвояемых углеводов и аминокислот, но и всех основных групп веществ: органических кислот, фенольных соединений, эфирных масел, азотистых и минеральных веществ.
Сахара. Они почти отсутствуют в сухих столовых винах, а остаточный сахар (1,5—2,5 г/л) состоит главным образом из несбраживаемых пентоз. В других типах вин в зависимости от степени сбраживания сахаров остается больше. Пектиновые вещества распадаются. Остаются несбраживаемые сахара (пентозы) и часть полисахаридов.
Органические кислоты. Определяя величину рН, они регулируют интенсивность и направленность биохимических реакций внутри дрожжевой клетки. В процессе брожения происходит также превращение кислот, как содержащихся в сусле, так и образованных при брожении. Дрожжи способны окислять уксусную, янтарную и лимонную кислоты по циклу ди- и трикарбоновых кислот.
Вместе с тем с помощью изотопа углерода доказано, что винные дрожжи в определенных условиях превращают пировиноградную кислоту в уксусную, а также синтезируют из аминокислот и углеводов янтарную, молочную, яблочную, глиоксалевую, фумаровую, галактуроновую и другие кислоты. Изменение титруемой кислотности определяется ее первоначальным значением в сусле. При высокой кислотности сусла (низкий рН) она снижается в процессе брожения, а при низкой кислотности — повышается.
Количество летучих кислот возрастает в анаэробных условиях и уменьшается при брожении сусла в присутствии кислорода. В процессе брожения выпадают в осадок кальциевые и кислые калиевые соли винной кислоты, растворимость которых уменьшается с повышением концентрации спирта.
Фенольные соединения. Они также претерпевают значительные изменения. В начале брожения, когда в сусле имеется кислород, фенольные вещества окисляются и дают продукты, обусловливающие покоричневение; часть хинонов, образуя димеры, выделяется из раствора. Во время брожения наблюдается восстановление хинонов за счет глютатиона дрожжей; цвет бродящего сусла или забродившей мезги светлеет.
Кроме того, фенольные вещества вступают в реакцию с белками и выпадают в осадок; образуют комплексные соединения с железом, ацетальдегидом, ингибируют микроорганизмы. В процессе брожения сусла на мезге в связи с повышением спиртуозности среды усиливается экстрагирование фенольных веществ, в том числе антоцианов и нефлавоноидных ароматических фенолокислот. Брожение сусла на мезге за счет фенольных веществ, придает особый аромат и вкус винам.
Минеральные вещества. При спиртовом брожении сусла они интенсивно потребляются дрожжами: примерно 20—30 % их переходит в дрожжевую массу. Часть минеральных веществ, связывает свободные кислоты сусла, увеличивая его нетитруемую кислотность или выпадая в осадок.
Содержание кальция может возрастать после обработки сусла мелом, при использовании плохо обработанных железобетонных резервуаров. Количество сульфатов может увеличиваться вследствие окисления сернистой кислоты, при обработке гипсом мезги или обработке бетонной поверхности резервуаров серной кислотой.
Микроэлементы. Их присутствие в виноградном сусле необходимо для питания дрожжей; они используются дрожжевыми клетками для синтеза витаминов и ферментов, участвуют в белковом обмене.
Азотистые вещества. Подвергаются при брожении сусла наибольшим изменениям. Дрожжевые клетки ассимилируют от 30 до 50 % имеющегося азота, в первую очередь легкоусвояемые формы: соли аммония и аминокислоты. В период брожения высокомолекулярная фракция азотистых веществ осаждается образующимся спиртом. Выпадают в осадок и танно-белковые комплексы.
Установлено, что при аэрации сусла потребление азотистых веществ дрожжами возрастает. С повышением температуры брожения накапливается биомасса дрожжей, увеличивается вынос азотистых веществ из сусла.
К концу брожения начинается отмирание части дрожжевых клеток и вслед за этим наступает их автолиз (разложение). При пониженных температурах автолиза (10—20°С) вино обогащается аминокислотами, витаминами, ферментами, липидами дрожжей, веществами аромата (эфиры, терпеноиды, жирные кислоты). При температурах, способствующих развитию бактерий (25—30°С), происходит бактериальное разложение азотистого комплекса отмерших дрожжей, аромат и вкус вина ухудшаются вплоть до появления посторонних тонов.
Ароматические вещества. В составе ароматических веществ сусла, насчитывающих более 300 наименований, при брожении происходит резкая перестройка состава.
Дрожжевые клетки на стадиях размножения и логарифмического роста потребляют терпеновые соединения, но выделяют набор высших алифатических и ароматических спиртов (3-фенилэтанол, тирозол) и сложных эфиров, изменяющих аромат сусла: вместо ярких терпеновых ароматов сорта появляются сильные тона розы, цветов липы, акации, меда, свойственные ароматическим спиртам и их сложным эфирам. Фруктово-цветочный запах имеют этиловые эфиры масляной кислоты и ее гомологов. Так формируются цветочные тона столовых вин.
При высоких температурах брожения, приятно пахнущие компоненты улетучиваются, а при хранении в условиях доступа кислорода воздуха — необратимо окисляются. Поэтому на стадии брожения сусла и формирования виноматериалов необходимо соблюдать постоянную герметизацию, удаляя избыток СО2.
На образование веществ аромата, букета вина положительно влияет смена рас дрожжей, что широко используется в классическом итальянском и французском виноделии.
Витамины и ферменты. Изменение витаминного состава сусла определяется расой дрожжей и условиями брожения. В первый период брожения большинство витаминов сусла поглощается дрожжами, затем дрожжевые клетки начинают сами синтезировать витамины биотин, никотинамид, пиридоксин и др. На заключительном этапе формирования виноматериалов и выдержке вина на дрожжах происходит переход витаминов в вино.
При брожении сусла инактивируются окислительные ферменты, а в стадии голодания дрожжи выделяют в среду протеиназу, β-фруктофуранозидазу и другие ферменты. Избыточная сульфитация сусла перед брожением, обработка бентонитом и другими сорбентами значительно обедняют вино полезными витаминами и ферментами.
Источники:
http://wine.historic.ru/books/item/f00/s00/z0000029/st086.shtml
http://znaytovar.ru/new85.html
http://vinocenter.ru/izmenenie-sostava-susla-v-processe-brozheniya.html





