
Репродуктивные органы – Фитогормоны виноградной лозы
Репродуктивные органы – Фитогормоны виноградной лозы
Словарь виноградаря
Нагрузка глазками – общее количество глазков, оставленных при обрезке на одном кусте или на 1 га виноградника.
Нагрузка побегами – количество вегетирующих побегов, оставляемых при обломке на кусте, на одном метре ряда или на 1 га виноградника.
Нагрузка урожаем —
- Количество соцветий или гроздей, оставляемых после обломки на одном кусте или плодовой стрелке.
- Урожай гроздей одного куста.
Неинфекционные болезни винограда – расстройство жизнедеятельности виноградного растения, обусловленное неблагоприятным влиянием внешних факторов среды.
Некорневое питание – питание растений удобрениями через наземные органы (главным образом через листья) путем опрыскивания растений водным раствором макро- и микроудобрений.
Некрозы винограда – специфическое заболевание винограда с общими симптомами отмирания отдельных участков тканей или частей растения.
Неукрывная культура винограда – способ выращивания винограда в открытом грунте без защитного укрытия кустов на зиму.
Нутация — круговые или качательные движения верхушек побегов, вызываемые более быстрым ростом то одной, то другой его стороны.
Обломка побегов – удаление части побегов с целью придания кусту необходимой формы, сохранения этой формы, установления оптимальной нагрузки побегами, оптимального соотношения между плодоносными и бесплодными побегами, а также необходимого соотношения между количеством побегов и урожаем на кусте.
Обрезка кустов винограда – ежегодное удаление части побегов и многолетних частей куста с целью придания ему определенной формы, поддержания этой формы (формировки), а также с целью установления оптимальной нагрузки побегами и гроздями с учетом силы роста куста и необходимости восстановления куста в случае повреждения его зимними морозами.
Одревеснение — пропитывание клеточных оболочек растений особым веществом – лигнином, придающим клеткам прочность и защищенность их от разрушительного действия бактерий.
Ожоги растений – местное повреждение тканей, вызываемое перегревом от солнца или при обработке химическими препаратами.
Оидиум – мучнистая роса, пепелица, грибное заболевание винограда, поражающее зеленые части куста на протяжении вегетации.
Окулировка — способ прививки, при котором на подвой прививают глазок привоя с частью древесины; прививка глазком (щитком).
Омолаживание — удаление старых многолетних ветвей (рукавов) и формирование из новых порослевых побегов или волчков отдельных рукавов или всей кроны куста (частичное или полное омолаживание).
Онтогенез растений – это совокупность закономерных морфологических и физиологических изменений, протекающих в организме от его зарождения из полового или из вегетативного зачатка до естественного отмирания, заложенного природой данного организма.
Операции с зелеными частями куста – комплекс технологических приемов, проводимых на виноградниках с зелеными частями куста в течение вегетации (обломка побегов, прищипывание, подвязка, чеканка, прореживание листьев и др.).
Опробкование (суберинизация) – отложение в толще клеточной оболочки растений особого вещества, суберина, делающего оболочку трудно проникающей для воды и газов.
Оптимальная нагрузка куста – число глазков и побегов, которое следует оставлять на растении при обрезке и затем при обломке части побегов для обеспечения хорошего развития куста и высокой его продуктивности.
Опыление – перенесение пыльцы с тычинок на рыльце пестиков цветков винограда (высших растений).
Опыливание – нанесение пылевидных ядохимикатов на поверхность растений с помощью опыливателей.
Органические удобрения – удобрения растительного или животного происхождения, содержащие азот и другие элементы питания растений преимущественно в форме органических соединений.
Подземный штамб – находящаяся в почве часть растения винограда, образовавшаяся из черенка, посаженного в школку или на постоянное место (часто неправильно называют корнештамбом).
Покой – состояние семян, почек или отдельных органов целого растения, во время которого у них прекращается видимый рост, но сохраняются скрытые процессы структурообразования.
Покой вынужденный – состояние растения, обусловленное неблагоприятными условиями среды. Наступает после физиологического покоя, когда виноград готов расти, но низкие температуры препятствуют его развитию.
Покой физиологический (органический) – определяется у виноградного растения состоянием зимующих почек, которые при повышении температуры не прорастают. При снижении температуры характеризуется глубоким изменением содержимого клеток и значительным снижением интенсивности обмена веществ во всем растении. Такое состояние почек у винограда длится с августа до декабря – января.
Полярность – свойственная растениям специфическая ориентация процессов и структур в пространстве, приводящая к возникновению морфо-физиологических градиентов и выражающаяся в различии свойств на противоположных концах или сторонах клеток, тканей, органов и всего растения.
Пористость почвы – суммарный объем всех пор, выраженный в процентах от общего объема почвы.
Порослевый побег – побег, развившийся на подземном штамбе куста из спящей почки. У винограда не бывает корневой поросли.
Почка – зачаток побега; орган нарастания, возобновления и вегетативного размножения растения; образуется на растущей части побега в пазухе листа.
Почковые вариации – стойкие модификационные или генотипические изменения, происходящие в отдельных почках и сохраняющиеся при вегетативном размножении в потомстве.
Прививка —
1. Хирургическая операция, при которой часть одного растения с почкой (глазком) соединяют с черенком, растущим побегом или штамбом другого растения для срастания.
2. Привитые растения, подготовленные для высадки в школку или на постоянное место.
Привитая культура винограда – способ возделывания европейских сортов винограда с использованием привитых саженцев, когда в качестве подвоя используются сорта, устойчивые к вредителям и другим неблагоприятным факторам.
Привитой – часть растения (черенок, глазок), которая переносится на другое растение (подвой) путем прививки; наземная часть привитого растения размещается выше места прививки.
Прищипывание побегов – декапитация, удаление верхушки побега (коронки), т.е. апикальной зоны с целью изменения передвижения ассимилянтов и питания к органам растения. Применяется для активизации роста пасынков, регулирования силы роста отдельных побегов, для ускорения вызревания лозы, для улучшения закладки эмбриональных и развития натуральных соцветий и лучшего их оплодотворения.
Пробковый камбий – вторичная образовательная ткань на побегах растений, образующая пробку и феллодерму.
Продуктивность виноградника – количество общей биомассы или массы гроздей, производимой виноградником за одну вегетацию; в случае гроздей продуктивность виноградника используется как синоним урожайности. Обобщающим показателем п. в. является коэффициент использования падающей ФАР – КПД ФАР виноградника.
Продуктивность побега (ПП) – количество сырой массы гроздей или массы сахара гроздей, создаваемой кустом в среднем на один развившийся побег (включая бесплодные); определяется как произведение коэффициента плодоношения на среднюю массу грозди куста. Является объективным критерием для оценки продуктивности сорта.
Продуктивность сорта – характеристика индивидуального растения, генетические свойства которого проявляются в агробиологических, морфологических и физиологических признаках (сила роста, плодоносность, облиственность, фотосинтез, дыхание и др.). Как генотипический признак, продуктивность сорта (сеянца, клона) характеризует способность растения производить определенную массу гроздей или сахара гроздей в расчете в среднем на один развившийся побег куста.
Прореживание гроздей – частичное удаление ягод в слишком плотных гроздях главным образом столовых сортов винограда с целью увеличения размеров оставшихся ягод.
Прореживание листьев – удаление затеняющих листьев в густых кронах для лучшего созревания гроздей винограда.
Прореживание соцветий – удаление отдельных соцветий или их частей с целью регулирования урожая винограда и его качества.
Простые минеральные удобрения – неорганические соединения, содержащие один из элементов питания растений: азот, фосфор, калий, магний и др.
Проуин – тонкий слой воскового налета, покрывающий стебли, листья и плоды винограда.
Пыльник – верхняя разросшаяся часть тычинки, содержащая пыльцу у цветковых растений.
Пыльца – совокупность пыльцевых зерен, образующихся в пыльниках семенных растений.
Расстилочная система – культура винограда без опор, при которой кусты стелются по земле.
Регенерация – способность растений восстанавливать удаленные органы и ткани, воспроизводить из отдельных частей новые растения.
Регрессия – количественные изменения одного признака в худшую сторону в зависимости от изменений другого.
Регуляторы роста – органические соединения, вызывающие стимуляцию или ингибирование процессов роста и развития растений.
Репродуктивные органы – органы, принимающие участие в размножении организма (у растений – соцветие, цветок, гроздь, семена).
Ретарданты – синтетические вещества разной химической природы, которые вызывают подавление роста стеблей и побегов, стимулируют плодоношение.
Рецессивность – форма взаимоотношений двух аллельных генов, при которой один из них – рецессивный – оказывает менее сильное влияние на соответствующие признаки особи, чем другой – доминантный; явление, противоположное доминантности.
Ризосфера – прикорневая зона, в которой сосредотачивается большое количество микроорганизмов, что связано с влиянием корневых выделений.
Ростовые вещества – фитогормоны, стимулирующие рост растений: ауксины, гиббереллины и др., а также некоторые природные соединения негормональной природы (фенолы, витамины) и др.
Фитогормоны виноградной лозы – Действие фитогормонов винограда и аналогичных им веществ
Содержание материала
Вегетативные органы
Ауксины и аналоги ауксинов. После обнадеживающих опытов Мu11еr- Stо11 (1939) в связи с воздействием ауксином на корнеобразование и развитие побегов виноградной лозы в целях производства привитых укорененных саженцев вновь были выдвинуты вопросы о корнеобразовании и аффинитете. Согласно исследованиям Тiziо (1967), даже незначительные концентрации индол-3-масляной кислоты (0,25—2,5 ppm) стимулируют корнеобразование легко окореняющегося сорта Мальбек, тогда как для труднее окореняющегося подвоя Кобера 5 ББ необходимы 0,25—25 ppm. В начале вегетационного периода, вследствие высокого содержания ингибиторов в побегах, возникает потребность в повышенных концентрациях ауксина; с увеличением эндогенных ростовых веществ и повышением способности к каллюсообразованию и корнеобразованию концентрацию применяемых ауксинов можно снизить (Тiziо, 1967). С целью улучшения окоренения зеленых черенков Turetskaya c сотр. (1966) предлагают 14-часовое погружение в 150 ppm индол-3-масляной кислоты. Это повышает процент окоренившихся черенков сорта Мадлен Анжевин с 70 до 100, а трудно окореняющегося сорта Тихоновский — с 4 до 74. По-видимому, генетически обусловленное более легкое окоренение некоторых сортов основывается на ускоренном и более интенсивном синтезе и активировании индол-3-уксусной кислоты у этих сортов (Turetskayac сотр., 1966). По мнению Eccher и Маrrо (1971), оптимальная концентрация наиболее часто используемых на практике растворов нафтил-3-уксусной кислоты зависит главным образом от величины черенков. Так, например, у длинных черенков высокие дозы (до 300 ppm нафтил-3-уксусной кислоты) могут стимулировать корнеобразование, тогда как небольшие дозы оказывают токсическое действие. Для видов Р. tricuspidata и Р. guinguefolia оптимальной для корнеобразования является концентрация 10- 6 до 10-4 (Lеrоuх, 1965). Shumann и Uhl (1975) усматривают в использовании метилэфира нафтил-3-уксусной кислоты и других стимулирующих рост и корнеобразование веществ технические преимущества для производства саженцев винограда. По-видимому, индуцированный индол-3- уксусной кислотой Rhizokalin (Ju11iаrd, 1966) в своем базипетальном передвижении не нуждается в мобилизации через посредство базипетально передвигающейся индол-3-уксусной кислоты (Ju11iаrd, 1967). По длине побегов создается корнеобразующий градиент, который не изменяется экзогенно под влиянием питательных и ростовых веществ. Очевидно, вместе с индол-3-масляной кислотой и гиббереллин (10-7—10-8М) оказывает стимулирующее воздействие на корнеобразование (Ju11iаrd, 1970а).
Погружение подвоев (V. solonis х V. Riparia 1616) на 24 h в концентрацию 50 ppm нафтил-3-уксусной кислоты непосредственно перед прививкой сорта Султанина определяет наибольший выход привитых саженцев, тогда как индол-3-уксусная кислота, индол-3-масляная кислота и гиббереллин оказали более слабое воздействие (Stinо с сотр., 1977). Если нижнюю треть подвоев прививок погрузить на 12 h в раствор ростовых веществ (0,1 % индол- 3-уксусной кислоты, 0,05% индол-3-масляной кислоты или 0,006% а-нафтил- 3-уксусной кислоты), повышается содержание питательных веществ в привое, а 90—95% привитых кустов вступают в плодоношение на третий год после посадки. Плодоносность глазков повышается, повышается и урожай винограда в первый год плодоношения на 7—92% (14—25 c/ha против 13 с/ha в контроле). Средний вес гроздей, однако, понижается (Khanin, Stepanova, 1970). По всей вероятности, лучшее срастание объясняется влиянием ауксинов на каллюсообразование (Fallot, 1970).
Как показывают опыты Sarkisova и Chailakhyan (1968) и Skene (1971), ауксин, по-видимому, участвует в метаболизме углеводов в черенках. Так, индол-3-уксусная кислота вызывает увеличение содержания сахаров и снижение концентрации крахмала (Skene, 1971). β-Индол-3-масляная кислота стимулирует физиологическую поляризацию черенков посредством быстрого смещения запасных веществ в сторону основания, где начинается их использование на корнеобразование; наконец, активизируется и ускоряется рост побегов (Sarkisova, Chailakhyan, 1968). Ессhеr и Маrrо (1971) и Skene также наблюдали стимулирование обмена веществ, повышенную активность камбия и повышенное потребление углеводов после обработки ауксином. Однако, наряду со стимулирующими рост свойствами, ауксины обладают и ингибирующим действием. Так, например, 200—400 ppm нафтил-3-уксусной кислоты задерживают прорастание почек примерно на 5 дней (Di Cesare, 1968), а обработка индол-3-уксусной кислотой черенков сорта Мерло, нарезанных незадолго до прорастания почек, замедляет этот процесс, если обрабатывается конус нарастания (Liuni, Poppi Struma, 1973a, b). Индол-3-масляная кислота также задерживает прорастание почек примерно на 7 дней (Sarkisova, Chailakhyan, 1968). Ауксины подавляют прорастание зимних почек не только непосредственно, а и косвенно. Исследования Alleweldt и Istar (1969) показывают, что 500, соответственно 1000 ppm индол-3-уксусной кислоты сокращают рост в длину пасынков на 38—56% у подвергнутых вершкованию растений, т.е. при устранении доминирующего влияния конуса нарастания.
Цитокинины. В отличие от ауксинов, применение кининов (1000 ppm бензиладенина) вызывает преждевременный выход почек из состояния покоя (W е a v е г с сотр., 1974а,Ь), что, по-видимому, обусловливается ускоренным распадом ингибиторов, наблюдаемым после применения бензиладенина (Wеavеr с сотр., 1968). Только у неокоренившихся черенков кинины (прежде всего рибозиды бензиладенина) стимулируют рост побегов. По мнению Pool и Powell (1975), это свидетельствует об участии образовавшихся в корнях цитокининов в развитии побегов. Редуцированный под влиянием индол-3-уксусной кислоты рост пасынков в длину отчасти восстанавливается при добавлении кинетина (100 и 1000 ppm) (Alleweldt, Istar, 1969). Очевидно, и результаты, полученные Scuflaire и Brebion (1967), объясняются задерживающим онтогенез действием цитокининов. С этой целью авторы погружали виноградные листья с черешками на 8 дней в раствор кинетина (10-2, 10-5, 10-7 М) в благоприятных условиях (освещение, температура, влажность воздуха). В этих листьях установлена стабилизация обмена веществ. Накопление продуктов фотосинтеза в контрольных листьях, как и распад существующих уже веществ, ограничивались под влиянием кинетина. Применение бензиладенина стимулирует движение ассимилятов из одного листа к другому, а также накопление 14С в физиологически более старых листьях в условиях затемнения (Quinlan, Weaver, 1969).
Гиббереллины. Многочисленные работы Alleweldt (1964, 1977; Alleweldt с сотр., 1972,1977) показывают, что рост побегов в длину подвержен влиянию в первую очередь гиббереллина, причем сорта V. vinifera заметно чувствительнее, чем американские. Вызванное фотопериодически коротким днем подавление роста побегов в длину преодолевается при помощи гиббереллина, а не в результате индуцированного коротким днем покоя почек (Аllеwе1dt, 1964). Наблюдавшееся подавление прорастания почек гиббереллином (Alleweldt, 1961) подтверждает Еris (1976 a,b). Применение гиббереллина подавляет прорастание почек во время глубокого вынужденного покоя (Е г i s, 1976а), следовательно, зимний покой становится более глубоким (Julliard, Balthazar d, 1965). Наряду с ростом побегов, обработка гиббереллином (1—100 ppm) уменьшает диаметр корней (Skene, Mullins, 1967). Подобно кининам, очевидно, и гиббереллины регулируют движение ассимилятов из одного побега к другому (Quin1аn, Мu11ins, 1970). Гиббереллин (10 ppm) поглощается как листьями, так и междоузлиями и передвигается к верхушке побега. Лучшие результаты в отношении поглощения гиббереллина получены на поверхности молодых листьев у их основания (Wеavеr и др., 1966а).
Абсциссины. Подобно индол-3-уксусной кислоте, абсциссиновая кислота подавляет прорастание зимних почек (Julliard, 1970b; Liuni, Poppi Struma, 1973a). Обработка базальных и апикальных почек дала одинаковые результаты, тогда как индол-3-уксусная кислота действует только на апикальные (Liuni, Poppi Struma, 1973b). Увеличение содержания абсциссиновой кислоты в зимних почках в фазе глубокого покоя приводит к выводу, что налицо специфическое участие абсциссиновой кислоты в эндогенном глубоком покое почек (During, Bachmann, 1975; During, Kisma1i, 1975). Независимо от возраста листьев, под влиянием 20 μΜ абсциссиновой кислоты повышается содержание антоцианов в листьях (Рiriе, Mullins, 1976). Обработка абсциссиновой кислотой пластинок молодых листьев стимулирует развитие соцветий и предотвращает отмирание зародышевых цветоножек. Через 24 h после обработки устанавливается известное подавляющее воздействие абсциссиновой кислоты на включение аминокислот в протеин (Mullins, Osborne, 1970). Инфильтрация абсциссиновой кислоты оказывает быстрое действие на срезанные листья: всего спустя 20 min, в зависимости от введенной концентрации, отмечается закрывание устьичных щелей, достигающее максимума через 70 min, после чего начинается частичное и повторное открывание (During, 1976b). Это свидетельствует о значительной функции абсциссиновой кислоты в регулировании движения устьиц. В противовес этому, абсциссиновая кислота не участвует в вызванном сменой света и темноты движении устьичных щелей, ибо ее содержание остается неизменившимся до и после включения, соответственно исключения света (During, 1976 а,b). Фотонастическое движение устьиц в значительно большей степени должно было бы определяться изменением межклеточной концентрации СО2, повышение которой в виноградных листьях ведет к закрыванию устьичных щелей, а не к изменению содержания абсциссиновой кислоты в листьях (Lоvеуs с сотр., 1973). Наряду с недостатком воды и кольцевание побегов приводит к повышению содержания абсциссиновой и фазеиновой кислот при условии, что отток ассимилятов из листьев понижен или полностью прекращен (Loveys, Kriedemann, 1974). Такое повышение содержания абсциссиновой кислоты, вызванное обогащением ассимилятами, приводит к закрыванию устьичных щелей, а при недостатке воды — и к улучшению водного потенциала листьев (During, 1978). Близкая к абсциссиновой фазеиновая кислота определяется как специфический ингибитор фотосинтеза in vivo (Kriedemann с сотр., 1975).
Аналоги этилена. Обработка этефоном (400 ppm) подавляет апикальный рост побегов в длину и задерживает прорастание почек примерно на 8—10 недель. При этом действие этефона значительно продолжительнее и равномернее, чем влияние повторного прищипывания (Lavее и др., 1977). Weaver и Роо1 (1971а) также установили после обработки 3000 ppm этефона замедленное прорастание почек на следующий год, а при обработке во время цветения сводилась на нет апикальная доминантность, т.е. начиналось образование пасынков и разделительной ткани на апикальных узлах.
Фитогормоны виноградной лозы
Содержание материала
X. Дюринг
ФИТОГОРМОНЫ ВИНОГРАДНОЙ ЛОЗЫ
ВВЕДЕНИЕ
Понятие фитогормоны охватывает 5 групп фитогенных веществ, передвигающихся в растении и в незначительных дозах индуцирующих или контролирующих различные физиологические процессы. Обозначенные как “химические сигналы”, гормоны в большей части обладают высокой подвижностью во всем растении, так что регулирующие функции этих веществ проявляются в месте их синтеза или, в известной степени при коррелятивных отношениях, в других органах. В более узком смысле к фитогормонам причисляются ауксины, гиббереллины, цитокинины, абсциссины и этилен. В разделе, где описываются выявляемые аналитическим путем фитогормоны виноградной лозы, речь идет и о дополнительных веществах, оказывающих ингибирующее действие, которые использовались в биологическом тексте. В следующем разделе рассматриваются вопросы действия синтезируемых аналогов фитогормонов и аналогичных им веществ, напоминающих фитогенные гормоны.
Исследовательская работа в целях определения гормонов виноградной лозы и их действия в последние годы разрослась до такой степени, что в объеме столь небольшой работы не представляется возможности охватить новейшую литературу по этому вопросу примерно после середины 60-х годов. Данные по этому вопросу можно найти в следующих обзорах: А11еwе1dt с сотр. (1967, а, b, 1975, 1977), Coombe (1973, 1976а), During,Allewе1dt (1974), During (1976a), Hidalgo, Candella (1973), а также в диссертациях Lott (1969), Hifnу (1971), During (1972), Ju1iiard (1973), Waitz (1975), Luckert (1976), Vollmer (1976), Koch (1977).
ЛОКАЛИЗАЦИЯ ФИТОГОРМОНОВ
Фитогормоны в вегетативных органах
Ауксины. Сезонная ритмика фаз роста и покоя виноградной лозы дает основание считать, что стимулирующие и подавляющие рост вещества участвуют в регулировании роста в отдельные времена года. Исследования Spiegel (1954) и Мashеvа (1975) показывают, что вещества, проявляющие ауксинную активность в биологическом тексте с побегами и почками, доказуемы, причем наиболее высокое их содержание установлено до распускания почек, после полного исчезновения растворимой в эфире ингибирующей фракции.
Эти результаты подтверждаются газохроматографическими анализами I s о d а (1975). Lilov и Nikolova (1974) установили, что физиологически молодые части побега, как и находящиеся под ними листья, отличаются большим содержанием ауксинов по сравнению с более старыми зонами побегов и листьями под ними. В главных и боковых корнях содержание ауксинов в течение вегетационного периода повышается, а после наступления зимнего покоя понижается. Наблюдавшийся Masheva (1975) базипетальный транспорт ауксинов свидетельствует в пользу предположения, что эти гормоны во время вегетационного периода передвигаются в корнях (Li1оv, Niко1оvа, 1974).
Цитокинины. Активность цитокининов в виноградной лозе была впервые доказана Nitsch и Nitsch (1965) в пасоке, а впоследствии, при применении того же метода, и в листьях (J а к о, 1970, 1977). V о 11 m е г (1976) подробно исследовал годичный ритм цитокининов в вегетативных органах разных сортов Vitis vinifera и на различных подвоях в условиях теплицы и открытого грунта. Низкое содержание их в зимних глазках в фазу эндогенного покоя сопровождается повышением активности в период после покоя до прорастания почек. До наступления сокодвижения это повышение, очевидно, объясняется синтезом или активацией цитокининов в почках, после чего наступает дополнительное накопление цитокининов, вероятно, из корней. Максимальное содержание цитокининов, установленное во время прорастания почек, сопровождается снижением его в молодых побегах до минимальных значений через 4—6 недель после прорастания. В июне содержание цитокининов вновь повышается в более старых листьях, тогда как в побегах и зимних глазках их содержание уменьшается с наступлением осени. Наподобие листьев, и в корнях установлено увеличение цитокинина во время вегетационного периода; более молодые части корней, однако, богаче цитокинином, чем более старые. Конус нарастания побегов также содержит больше цитокинина, чем листья. У всех исследовавшихся сортов и сочетаний прививок установлено повышение концентрации цитокинина по мере усиления интенсивности сокодвижения до прорастания почек и понижение — с ослаблением интенсивности плача после распускания почек. Очевидно, концентрация цитокинина в пасоке определяется преимущественно привоем.
Исследования Lilov и Andonowa(1976) подтверждают эти результаты. Во время роста ягод наступает снижение содержания цитокининов в листьях, а во время их созревания оно вновь повышается. Примерно до июля в побегах и корнях устанавливается повышение цитокинина, после чего его активность начинает снижаться до середины ноября, а затем опять повышается в декабре. Итак, за исключением листьев, во время эндогенной фазы покоя во всех органах виноградной лозы устанавливается незначительная, а впоследствии в различной степени возрастающая активность цитокининов.
Исследования в связи с повышением содержания цитокининов при различной температуре показали, что хранение в условиях низкой температуры (+1°С) повышает активность цитокининов в вызревших побегах (Skene, 1972), а также, что разная температура питательного раствора приводит к изменению содержания отдельных цитокининовых фракций в корневом экссудате (Skene и Keridge, 1967). Нарушение соотношения источник — потребитель (source — sink) у плодоносящих кустов путем удаления гроздей понижает активность фотосинтеза и в то же время усиливает образование цитокининовых глюкозидов в листьях (Hoad с сотр., 1977). Повышение содержания цитокинина примерно в 20 раз установлено в пасоке, когда в питательный раствор добавляли хлорхолинхлорид (ССС) (Skene, 1968, 1970).
Наоборот, снижение активности цитокинина в корнях отмечалось при уменьшении доз N или К. В условиях без удобрения NPK в корнях не обнаруживалось цитокининов (Jако, 1974).
Гиббереллины. До прорастания почек Isоdа (1973) установил повышение активности гиббереллинов (GA) в почках. В корневом экссудате и в экстрактах из листьев и корней Skene (1967) обнаружил вещества, Rf- значения и биологическая активность которых соответствовали GA4 и GA7. В усиках установлено большее содержание гиббереллина, чем в других органах побега; закручивание усиков сопровождается ростом гиббереллинов (Маnanкоv, 1976). По мнению Kocherzhenko и Maiko (1967), между содержанием гиббереллина в листьях и ростом существует тесная корреляция. Так, например, в условиях продолжительного освещения и длинного дня в листьях быстрорастущих растений при помощи теста прорастания гороха установлены гиббереллины, которых не обнаружено в условиях короткого дня при редуцированном росте. Удаление гроздей, по данным Hoad с сотр. (1977), ведет к уменьшению гиббереллиноподобных веществ в листьях.
Абсциссины и ингибиторы. Впервые в пасоке виноградной лозы при помощи биологического теста Rapp и Ziegler (1971)установили абсциссиновую кислоту на основе Rf-значений в различных жидкостях, поведения в ультрафиолетовой области, а также инфракрасного и массового спектров. На основе этих методов впоследствии были исследованы сезонные изменения абсциссиновой кислоты в побегах и листьях: содержание абсциссиновой кислоты в листьях, где она синтезируется, не претерпевает существенных изменений в течение вегетационного периода. В побегах, наоборот, обнаруживается четкая сезонная динамика, причем максимум устанавливается в эндогенную фазу покоя (During, Alleweldt, 1973; Cheng с сотр., 1974). Решающим фактором повышения содержания абсциссиновой кислоты можно считать уменьшение длины дня, что, однако, не снижало ее количества в листьях, подвергнутых раздражению фотопериодом. Взамен этого, уменьшение длины дня увеличивает абсциссиновую кислоту в побегах, что свидетельствует о ее передвижении из листьев к побегам (Alleweldt, During, 1972). В апикальных частях побегов нередко устанавливалось большее содержание абсциссиновой кислоты, чем в секторах у основания (Lilov, Angelova, 1977а; During, неопубликованные данные). Как показывают опыты Lоveys и Kriedemann (1973), наблюдается быстрая и четкая реакция содержания абсциссиновой кислоты в листьях на продолжительный водный дефицит; при пониженной интенсивности транспирации и фотосинтеза, а также при повышении водного потенциала, содержание абсциссиновой кислоты в листьях увеличивается, тогда как после повторного полива наблюдается обратное. Наряду с повышением содержания абсциссиновой кислоты в листьях увядающих виноградных кустов устанавливается еще большее его повышение в побегах (During, Sсiеnz а, 1975), что, по всей вероятности, приводит к прекращению роста побегов в длину. Исследования показали, что суточный ход транспирации в большинстве случаев находится в отрицательной корреляции с содержанием абсциссиновой кислоты в листьях (During, Scienza, 1975). Из листьев выделена также близкая абсциссиновой фазеиновая кислота (Loveys, Kriedemann, 1974).
В нейтральных, основных и кислых экстрактах, из побегов Тiziо с сотр. (1968) установили при помощи биологического теста ингибиторы. Weaver с сотр. (1968) и Sarkisova с сотр. (1976) установили в начале периода покоя повышение феноловых ингибиторов в побегах и почках, содержание которых уменьшается с началом вегетации. Это эндогенное содержание ингибиторов непосредственно связано с морозостойкостью отдельных сортов (Sarkisova с сотр., 1976).
Фитогормоны в репродуктивных органах
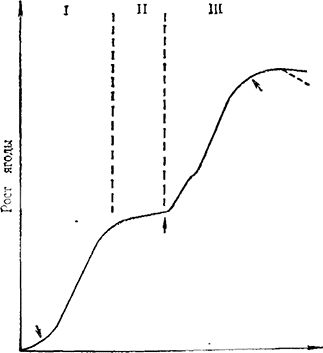
Рис. 1. Динамика содержания ауксинов во время роста ягоды винограда
I — цветение; II — начало созревания; III — физиологическая зрелость
Ауксины. Впервые ауксиноподобные вещества в ягодах винограда установили Nitsсh с сотр. (1960) и Сооrubе (1960). При помощи теста A vena mesocotyle установлено повышение содержания ауксиноподобных веществ в I фазе роста ягод (рис. 1) и понижение его при временном приостановлении роста во II фазе. Исследования Alleweldt и Hifny (1972) и Farmahan и Pandey (1976) подтверждают этот ход ауксиноподобных веществ.
Ауксин в незначительной концентрации доказан в ягодах при помощи газовой хроматографии (Сооmbе, Hale, 1973) и хроматографии с использованием жидкости под большим давлением (During, 1977). Применение гиббереллина калия повысило уже через 3 h после обработки активность ауксинов в ягодах; этот повышенный уровень ауксинов вновь понижается во II фазе роста ягод (Bertrand, Weaver, 1972а). Во время стратификации активность ауксинов в семенах повышается, достигая максимума при прорастании (Сhоhan, Dhi11оn, 1976; Касhru с сотр., 1969).
Цитокинины. Новейшие исследования при помощи теста в каллюсе сои показывают, что неидентифицированные до сих пор активные вещества (Waitz, 1975; Сhаско с сотр., 1976; Lilov, Andonova, 1976) появляются в ягодах преимущественно в начале их роста. Относительно содержания цитокининов в отдельные фазы роста имеется немного данных, которые, однако, совпадают. Waitz (1975) установил совсем незначительные концентрации цитокинина в соцветиях в полном цветении, в то время как Lilov и Andonova в этой фазе обнаружили большие концентрации, чем в развивающихся впоследствии ягодах. Пробы в исследованиях брали в один, соответственно в два срока. В разгар цветения и во время образования ягод Lilov и Andonova обнаружили незначительную концентрацию цитокининов, а и Waitz сообщает о цитокининовом минимуме. В противовес этому Сhаско с сотр. (1976) установили максимальные значения, уменьшающиеся в фазу интенсивного роста (I фазу).
Если пренебречь наблюдаемым в III фазе повторным повышением содержания цитокинина, то можно принять, что сделанные констатации о появлении цитокининов в ягодах преимущественно в первую фазу усиленного роста в значительной мере совпадают.
Гиббереллины. Наряду с ауксинами Сооmbе (1960) впервые установил гиббереллиновую активность в незрелых ягодах многих сортов. С началом нарастания ягод Iwahоri с сотр. (1968), Itо с сотр. (1969), а также Lilον и Christov (1977) обнаружили максимальное содержание гиббереллина, задерживающие на одном уровне около двух недель, а затем понижающееся. До сих пор повышение содержания гиббереллиноподобных веществ в III фазе созревания было установлено только Farmahan и Pandey (1976). Бессемянные сорта почти без исключения отличаются меньшей активностью гиббереллина по сравнению с сортами с семенами (Сhai1akhуаn, Sarkisova, 1965; Weaver, Pool, 1965а,b; Lilov, Christov, 1977; Nazemi11e, 1977), что выражается несколько замедленным нарастанием незрелых ягод с семенами (Iwahori с сотр., 1968). Незначительное содержание гиббереллина в незрелых ягодах соответствует его меньшим значениям также в листьях, соцветиях и гребнях, тогда как у сортов с семенами в генеративных органах установлено большое содержание гиббереллина, чем в вегетативных (С hailakhyan, Sarkisova, 1965). Weaver и Pool (1965а) обнаружили активность гиббереллиноподобных веществ до оплодотворения в узлах, несущих соцветия. Кольцевание вызывает изменения в спектре гиббереллинов, что означает, что содержание гиббереллина в кислой фракции повысилось, а в нейтральной — понизилось (Weaver, Pool, 1965 b, с).
Абсциссины. О локализации абсциссиновой кислоты в ягодах винограда сообщают Coombe и Hale (1973), Inаbа с сотр. (1976) и During (1973, 1974, 1977), которым при помощи газовой хроматографии, соответственно хроматографии в жидкости под высоким давлением, удалось установить увеличение этого гормона с началом созревания ягод и понижение — перед окончательным созреванием. During (1974) установил почти параллельное повышение абсциссиновой кислоты в мякоти ягод и семенах, в то время как Coombe и Hale (1973) не удалось установить увеличения абсциссиновой кислоты в семенах; Lilov и Angelova (1977b) и Scienza (личные сообщения) также обнаружили лишь незначительное количество абсциссиновой кислоты в семенах. Lott (1968, 1969), однако, установил в выделенных семенах винограда высокое содержание абсциссиновой кислоты, которое уменьшается во время стратификации параллельно повышению степени их готовности к прорастанию. Констатации (неопубликованные) During и данные Chоhan и Dhil1 on (1976) подтверждают эту зависимость. По мнению Coombe (1976a,b) содержание абсциссиновой кислоты повышается примерно в течение 10 недель после цветения, причем в кожице ягод увеличение наступает раньше и быстрее, чем в мякоти.
Этилен. Подобно неклимактерическим плодам черешни (не претерпевающим резких изменений в дыхании во время созревания), в ягодах винограда были установлены совсем незначительные концентрации этилена, во время созревания проявляющие тенденцию к снижению (Coombe, На1е, 1973). В новейших исследованиях Inаbа с сотр. (1976), Косh (1977) и А11еwе1dt и Koch (1977) также установлено внутриклеточное содержание этилена. Независимо от относительного значения (на ягоду или грамм свежего веса), в начале II фазы установлено повышение содержания этилена, которое, однако, перед началом созревания вновь быстро понижается.
Источники:
http://dom-sad-og.ru/slovar-vinogradarya/
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/fitogormony-vinogradnoy-lozy-2.html
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/fitogormony-vinogradnoy-lozy.html





