
Превращение и передвижение ассимилятов – Питание виноградного растения
Превращение и передвижение ассимилятов – Питание виноградного растения
научная статья по теме ОСОБЕННОСТИ СУТОЧНОГО РИТМА ПЕРЕДВИЖЕНИЯ АССИМИЛЯТОВ В ВИНОГРАДНОЙ ЛОЗЕ Биология

Цена:
Авторы работы:
ЧАНИШВИЛИ ШОТА ШАЛВОВИЧ
Научный журнал:
Год выхода:
Текст научной статьи на тему «ОСОБЕННОСТИ СУТОЧНОГО РИТМА ПЕРЕДВИЖЕНИЯ АССИМИЛЯТОВ В ВИНОГРАДНОЙ ЛОЗЕ»
ФИЗИОЛОГИЯ РАСТЕНИЙ, 2004, том 51, № 1, с. 57-61
ОСОБЕННОСТИ СУТОЧНОГО РИТМА ПЕРЕДВИЖЕНИЯ АССИМИЛЯТОВ В ВИНОГРАДНОЙ ЛОЗЕ
© 2004 г. Ш. Ш. Чанишвили, М. Д. Долидзе, Г. Ш. Бадридзе, Т. Ф. Барблишвили
Институт ботаники им. Н. Кецховели Академии наук Грузии, Тбилиси Поступила в редакцию 22.01.2002 г.
Изучены особенности оттока фотоассимилятов из листа виноградной лозы (УШя у1т/ега Ь., уаг. Як^НеИ) в естественных условиях и под влиянием затемнения, дефолиации и АБК при коротких экспозициях (2, 6, 24 ч), а также дневная динамика содержания крахмала в листьях. Транспорт асси-милятов из листа виноградной лозы происходил в основном ночью и начинался после 20:00-21:00. Однако такая периодичность транспорта не была строго детерминирована и могла меняться под воздействием ряда факторов. Затемнение, дефолиация и обработка АБК инициировали транспорт фотоассимилятов из листьев в дневные часы. Затемнение и АБК оказались более эффективными по сравнению с дефолиацией. Затемнение и дефолиация также существенно меняли характер распределения фотоассимилятов в стебле. Подтверждено мнение, что и в виноградной лозе отток ас-симилятов из донорного листа регулируется механизмом обратной связи и, по всей вероятности, сильно зависит от уровня углеводов.
УШз уш/ега – транспорт фотоассимилятов – затемнение – дефолиация – АБК – крахмал
Ритм – это внутреннее свойство организма, обусловленное периодичностью смены активности и депрессии ферментов [1]. Согласно литературным данным, процессы синтеза и гидролиза проявляют определенную дневную и суточную периодичность. Установлено, что в обычных условиях интенсивные биосинтетические процессы, наблюдаемые в дневные часы, ночью сменяются интенсивными гидролитическими процессами, когда в растении более активно идет передвижение веществ. Определенная ритмичность выявлена в оттоке фотоассимилятов из листа, который в течение суток происходит неравномерно и характеризуется видовой специфичностью [2; 3, с. 462-465].
Неравномерный отток ассимилятов из листа еще не нашел четкого объяснения. Существует предположение, что процесс транспорта обусловлен типом углеводов, которые аккумулируются в листьях. Так, при накоплении в листьях крахмала транспорт начинается ближе к концу фотопериода, в отличие от накопления дисахаридов [4].
Имеются указания на влияние интенсивности освещения на транспорт ассимилятов, но здесь трудно сделать однозначный вывод. Часть авторов считает, что освещение стимулирует транспорт веществ, другими же установлено, что этот
Адрес для корреспонденции: Чанишвили Шота Шалвович. 380007 Грузия, Тбилиси, Коджорское шоссе, 1. Институт ботаники АН Грузии. Электронная почта: gbadridze@yahoo.сom
процесс не зависит от освещения или же более интенсивно идет в темноте [3, с. 462-465].
В ранних работах мы установили, что для виноградного растения характерен своеобразный суточный ритм передвижения ассимилятов: при нормальных условиях оно приостановлено днем и начинается лишь вечером [2]. Целью настоящей работы являлось определение момента начала оттока фотоассимилятов из листа и степени стабильности этого процесса, а также выявление регулирующих его механизмов.
Объектом исследований служили однолетние побеги плодоносящих лоз винограда (Уitis уМ-/ега Ь., уаг. Як^ИеН). Лозы были сформированы по двусторонней шпалерной системе. По обе стороны куста на плодоношение оставляли 7-9 побегов. На побеге было по 20-25 вполне сформировавшихся листьев, со средней площадью 85-95 см2 каждый. Метку вводили в один интактный лист отдельного побега [5, с. 66-75]. Для устранения влияния других частей куста базальную часть побега окольцовывали сразу после введения метки. Опыты проводили в период интенсивного роста ягод.
С целью установления начала оттока фотоассимилятов из листа для каждой экспозиции брали отдельный побег и метку вводили в лист с таким же пластохронным индексом, что и для других экспозиций. В фотосинтетической камере лист
Таблица 1. Дневная динамика оттока фотоассимилятов из подкормленного 14С02 листа виноградной лозы
Время после введения метки,ч
Время введения CO
радиоактивность, тыс. имп./(мин г сухой массы)
листа черешка листа черешка листа черешка листа черешка
0 140 ± 7 0 72 ± 2 0 148 ± 7 0 47 ± 2 0.20 ± 0.06
3 135 ± 6 0 87 ± 3 0 158 ± 6 0.40 ± 0.01 – –
8 148 ± 6 0 66 ± 2 0.30 ± 0.01 – – – –
11 158 ± 6 0.80 ± 0.02 – – – – – –
Примечание. Прочерк – данные не приводятся.
выдерживали 5 мин, радиоактивность составляла 1.5 МБк/л воздуха, концентрация С02 – 1%. После введения 14С02 лист оставляли на побеге в течение заданной экспозиции, после чего его срезали с черешком и фиксировали при 105°С. С помощью торцевого счетчика Т-25 БФЛ (радиометр ПП-8, изготовлен в ТСХА, Москва) определяли радиоактивность черешка. Обнаружение в нем радиоактивности служило сигналом о начале оттока фотоасси-милятов из листа.
В серии экспериментов при изучении интенсивности оттока фотоассимилятов и их распределения между органами побега интактный лист подкармливали 14С02 и оставляли на побеге в течение заданного периода (4, 6 или 24 ч), после чего побег фиксировали. Суммарный уровень радиоактивности всех органов принимали за 100%. Сравнивая радиоактивность других органов (кроме подкормленного листа) с этой величиной, вычисляли долю экспортированной 14С ассимиля-тов, принимая ее за показатель интенсивности оттока. Распределение фотоассимилятов между органами побега оценивали по проценту радиоактивности отдельных органов от величины радиоактивности экспортированных из листа-донора 14С ассимилятов.
Для изучения влияния дефолиации проводили 75%-ную дефолиацию побега, оставляя только 45 листьев среднего яруса. Это были завершившие рост листья, которые являются основными донорами, снабжающими верхушку побега и гроздь фотоассимилятами. В один из оставшихся листьев вводили 14С02. После заданной экспозиции (4, 6, 24 ч) побег фиксировали (105°С). По распределению радиоактивности в междоузлиях судили о транспорте ассимилятов.
В экспериментах с затемнением, лист (первый или второй выше грозди) затемняли фольгой и спустя 4, 6 или 24 ч побег фиксировали. За транспортом следили по распределению радиоактивности в побеге.
В опытах с АБК лист из зоны грозди дважды на 1 мин погружали в раствор этого вещества (40 мг/л) (“Sigma Chemicals Co.”, США). Через 4, 6 или 10 ч побег фиксировали для изучения транспорта.
Количество крахмала в листьях определяли колориметрически по методу Починка [6, с. 134138]. Для этого 0.2 г сухого, измельченного материала заливали 80%-ным Ca(NO3)2, тщательно гомогенизировали и центрифугировали (10002000 g, 7-10 мин). В центрифугат, содержащий крахмал, добавляли йод. Полученное темно-синее соединение йода с крахмалом растворяли в 10 мл 0.1 N NaOH и после добавления 0.3 мл 0.5%-ного йода и 2 мл 1 N HCl измеряли оптическую плотность раствора при длине волны 580-610 нм на спектофотоколориметре SPEKOL11(“Carl Zeiss”, Германия).
В таблицах представлены средние величины трех биологических повторностей и их стандартные отклонения.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изучение дневной динамики передвижения фотоассимилятов показало, что транспорт веществ из листьев в виноградной лозе начинался в конце дня после 20:00-21:00 (табл. 1). Так как транспортируемые вещества должны пройти черешок, прежде чем окажутся в стебле, появление метки в черешке служит четким свидетельством начала процесса оттока. Отчетливо видно, что чем раньше метка включается в состав фотоассимилятов (в том числе и крахмала), тем позже она оказывается в транспортируемых веществах (т.е. в сахарозе). Это объясняется тем, что в виноградной лозе в течение дня происходит накопление крахмала в листе, а к вечеру начинается его гидролиз, и углеводы транспортируются из листа в стебель в виде сахарозы. Регуляция транспорта должна происходить на генном уровне, с помощью определенных механизмов с обратной связью [7].
Таблица 2. Влияние затемнения на начало оттока фотоассимилятов из подкормленного 14С02 листа виноградной лозы
Продолжительность затемнения, ч Время введения 14С02
9:00 13:00 18:00 20:00
радиоактивность, тыс. имп./(мин г сухой массы)
листа черешка листа черешка листа черешка листа черешка
2 217 ± 9 0 98 ± 4 0 180 ± 5 0 37 ± 1 0.30 ± 0.01
3 239 ± 12 0 128 ± 5 0.70 ± 0.02 – – – –
4 200 ± 8 0.90 ± 0.02 – – – – – –
Примечание. Прочерк – данные не приводятся.
Таблица 3. Влияние затемнения, дефолиации и АБК на отток фотоассимилятов из подкормленного 14С02 листа в виноградной лозе
Вариант Продолжительность воздействия, ч Интенсивность оттока, % от суммарной радиоактивности на момент определения Распределение фотоассимилятов, % от экспортированной радиоактивности
верхняя часть побега нижняя часть побега гроздь
Контроль 4 0 0 0 0
24 15.4 0 82.0 18.0
Затемнение 4 16.4 32.7 67.3 0
6 20.4 26.6 45.4 28.0
Контроль 4 0 0 0 0
24 40.8 0 44.3 55.7
Дефолиация 4 0 0 0 0
24 55.0 38.1 51.9 10.0
Дефолиация + АБК 6 12.4 26.0 74.0 0
24 58.7 18.9 31.3 49.8
При накоплении крахмала до определенного уровня “включаются” процессы его гидролиза. В пользу этого соображения свидетельствуют опыты с затемнением листа.
При изучении влияния затемнения на транспорт веществ было выявлено, что оно ускоряет начало транспорта из листьев (табл. 2). Время начала транспорта зависит от времени начала затемнения листьев: в утренние часы требуется 4 ч затемнения, в полдень – 3 ч, а вечером и двух часов достаточно для того, чтобы из листа начался отток фотоассимилятов. Роль метаболитов-регуляторов в таком случае должны выполнять углеводы. Это вполне вероятно, так как углеводы -основные вещества, образующиеся в ходе фотосинтеза, и к тому же известно, что они принимают активное участие в экспрессии генов у растений [8].
Таким образом, затемнение, имитируя ночные условия, вызывает включение внеочередных механизмов, стимулируя отток.
Интересно, что затемнение меняет направление транспорта (табл. 3). Если обычно в этот период вегетации фотоассимиляты в
Для дальнейшего прочтения статьи необходимо приобрести полный текст. Статьи высылаются в формате PDF на указанную при оплате почту. Время доставки составляет менее 10 минут. Стоимость одной статьи — 150 рублей.
Пoхожие научные работы по теме «Биология»
БАКИРОВА Г.Г., ЧИКОВ ВЛАДИМИР ИВАНОВИЧ — 2004 г.
Питание растений
Процесс поглощения и усвоения растениями из окружающей среды химических элементов, необходимых для их жизни; заключается в перемещении веществ из среды в цитоплазму растительных клеток и их химическом превращении в соединения, свойственные данному виду растений. Поглощение и усвоение питательных веществ (анаболизм) вместе с их распадом и выделением (катаболизм) составляют Обмен веществ (метаболизм) — основу жизнедеятельности организма.
В составе растений обнаружены почти все существующие на Земле химические элементы. Однако для П. р. необходимы лишь следующие: углерод (С), кислород (О), водород (Н), азот (N), фосфор (Р), сера (S), калий (К), кальций (Ca), магний (Mg), железо (Fe) и Микроэлементы: бор (В), марганец (Mn), цинк (Pb), медь (Cu), молибден (Mo) и др. Элементы питания поглощаются из воздуха — в форме углекислого газа (CO2) и из почвы — в форме воды (H2O) и ионов минеральных солей. У высших наземных растений различают воздушное, или листовое, питание (см. Фотосинтез) и почвенное, или корневое, питание (см. Минеральное питание растений). Низшие растения (бактерии, грибы, водоросли) поглощают CO2, H2O и соли всей поверхностью тела.
Потребность растительного организма в различных элементах неодинакова; наибольшая — в кислороде и водороде. Это объясняется тем, что живое растение на 80—90% состоит из воды, т. е. из кислорода и водорода в отношении 8: 1. Кроме того, растение расходует за свою жизнь в процессе транспирации (См. Транспирация) в сотни раз больше воды, чем его собственная масса (для предотвращения перегрева). Основу сухого вещества растения наряду с углеродом (45%) составляют также кислород (42%) и водород (6—7%). На долю элементов минерального питания, среди которых преобладают азот и калий, приходится всего 5—7% сухого вещества растения. Ни один элемент питания не может быть заменен другим (так называемый принцип незаменимости питательных элементов). Отсутствие или большой недостаток любого из них неизбежно приводит к прекращению роста и к гибели растения. Каждый из элементов выполняет в растительных тканях свою уникальную функцию, неразрывно связанную со всеми др. отправлениями организма. Так, углерод вместе с водородом и кислородом составляет основу всех молекул органических соединений (см. Биогенные элементы). Вещества, состоящие только из этих трёх элементов (углеводы),— главный субстрат дыхания (См. Дыхание). Из полимерных углеводов состоят также оболочки растительных клеток. Каждый вид и даже сорт растений поглощает преимущественно те элементы, которые в наибольших количествах нужны для свойственного ему обмена веществ. Поэтому, например, содержание калия в растениях обычно в десятки раз превышает содержание натрия, хотя в почвах отношение между этими элементами обратное. Некоторые виды растений способны накапливать в своих тканях редкие элементы (например, лантан), чем пользуются при геологической разведке (см. Индикаторные растения).
Типы питания. В зависимости от источника поглощаемого углерода различают несколько типов П. р. Часть низших растений (все грибы и большая часть бактерий) может использовать углерод только из органических соединений, в которых он содержится в восстановленной форме. При окислении таких соединений в процессе дыхания освобождается запасённая в них химическая энергия, которая затем может расходоваться на различные эндергонические (т. е. требующие затрат энергии) процессы: синтез более сложных соединений, передвижение веществ в растении и др. Питание этого типа называется гетеротрофным, а растения, потребляющие органические источники углерода,— гетеротрофными (см. Гетеротрофные организмы); питание за счёт мёртвых органических остатков называется сапрофитным, а растения, питающиеся мёртвыми органическими остатками,— сапрофитами (См. Сапрофиты). Этот тип питания свойствен всем гнилостным грибам и бактериям. Гетеротрофы, живущие за счёт органических соединений др. живых организмов, называются паразитами (См. Паразиты). К ним относятся все грибы и бактерии — возбудители болезней животных и растений, а также некоторые высшие растения, например заразиха, высасывающая с помощью специальных присосок соки др. растений. Паразитическое П. р. отличается от Симбиоза, при котором происходит постоянный обмен продуктами жизнедеятельности, полезный для обоих партнёров. Симбиотический П. р. наблюдается, например, у азотфиксирующих бактерий, поселяющихся в клубеньках на корнях бобовых растений (см. Азотфиксация), у шляпочных грибов, гифы которых проникают в корневые ткани древесных растений (см. Микориза), а также у лишайников, представляющих собой группу грибов, находящихся в постоянном сожительстве с водорослями. Большая часть растений способна усваивать углерод из углекислого газа, восстанавливая его до органических соединений. Этот тип питания называется автотрофным (см. Автотрофные организмы). Он свойствен всем высшим зелёным растениям, а также водорослям, некоторым бактериям. Восстановление CO2 до органических соединений требует затрат энергии либо за счёт поглощаемого солнечного света (фотосинтетики), либо за счёт окисления восстановленных соединений, поглощаемых из внешней среды (хемосинтетики).
Благодаря П. р. осуществляется большой биогеохимический Круговорот веществ в природе (рис. 1). Автотрофные (главным образом зелёные, или фотосинтезирующие) растения дают начало этому круговороту, удаляя из атмосферы CO2 и создавая богатые химической энергией органические вещества. Гетеротрофные растения (главным образом сапрофиты) замыкают этот круговорот, разлагая мёртвые органические остатки до исходных минеральных веществ.
В процессе фотосинтеза растения не только поглощают вещества, но и накапливают энергию. Один из первичных продуктов фотосинтеза — сахара. При соединении 6 грамм-молекул CO2 и такого же количества H2O образуется 1 грамм-молекула глюкозы (180 г). Этот процесс происходит с поглощением 674 ккал (1 ккал = 4,19 кдж) энергии солнечного света, которая и запасается в химических связях сахара. Вместе с молекулами сахара эта запасённая химическая энергия может затем переместиться в другие, нефотосинтезирующие части растений, например в корень. Здесь в процессе дыхания она может освобождаться для синтеза более сложных соединений и для др. процессов жизнедеятельности растительных клеток. Хотя в фотосинтезе непосредственно участвуют только CO2 и H2O, для его осуществления и в особенности для последующих превращений его первичных продуктов необходимы все др. элементы П. р., в каких бы ничтожных количествах они не содержались в растении.
Превращения питательных веществ происходят в различных органах и тканях и связаны друг с другом в непрерывный круговорот веществ в растительном организме (рис. 2). В листьях в процессе фотосинтеза из CO2 воздуха и поступающей из корня H2O образуются первичные органические продукты (ассимиляты). Один из них — сахароза — универсальная форма транспортировки углевода. Из фотосинтезирующих клеток листа сахароза поступает в специальную транспортную систему — ситовидные трубки флоэмы (См. Флоэма), обеспечивающие нисходящее перемещение веществ сначала по листовым жилкам, а затем по проводящим пучкам стебля в корень. Здесь ассимиляты покидают ситовидные трубки и распространяются по тканям корня. Навстречу притекающим из листьев ассимилятам движутся вода и ионы минеральных солей, которые сначала связываются поверхностью корневых клеток, а затем через клеточную мембрану проникают внутрь клеток. При этом одни элементы (калий, натрий, в значительной степени кальций, магний и др.) поступают в пасоку (См. Пасока) и подаются в надземные органы в неизменном состоянии. Другие (например, азот), встречаясь с центробежным потоком ассимилятов, вступают с ним во взаимодействие, включаясь в состав органических соединений (аминокислот и амидов), и в таком измененном виде поступают в пасоку. Наконец, третьи (такие, как фосфор), проходя через ткани корня, также включаются в органические соединения (нуклеотиды, фосфорные эфиры сахаров), но затем, снова отщепляясь, поступают в пасоку главным образом в виде свободных ионов. Так или иначе элементы корневого П. р. вместе с водой поступают в сосуды ксилемы (См. Ксилема) — вторую транспортную систему растения, обеспечивающую восходящее перемещение веществ в надземные органы. Движение воды и растворённых в ней веществ по сосудам происходит за счёт корневого давления и транспирации. В листе эти вещества из сосудов проникают в фотосинтезирующие клетки, где происходит их вторичное взаимодействие с ассимилятами. При этом образуются разнообразнейшие органические и органо-минеральные соединения, из которых после ряда усложнений развиваются новые органы растения.
Роль питания. П. р. обеспечивает веществами и энергией следующие процессы: поддержание жизнедеятельности (возмещение убыли питательных веществ при дыхании и выделении в наружную среду), рост органов, отложение веществ в запас и, наконец, воспроизведение потомства (образование плодов и семян). При недостаточном П. р. питательными веществами обеспечиваются в первую очередь процессы, связанные с жизнедеятельностью и воспроизведением потомства. При умеренном недостатке П. р. рост молодых частей растения (верхних листьев, корневых окончаний) ещё продолжается за счёт реутилизации, т. е. повторного использования питательных элементов путём их оттока из более старых листьев. При резком недостатке П. р. рост прекращается, и все питательные ресурсы направляются на главную функцию растительного организма — воспроизведение потомства. В этих условиях ячмень, например, имеет высоту всего 4—5 см, но образует 2—3 вполне нормальные зерновки. Избыток тех или иных элементов П. р. так же вреден, как и их недостаток.
Создание наилучших условий почвенного П. р. путём орошения и внесения удобрений — наиболее эффективное средство управления урожаем с.-х. растений. В закрытом грунте (парники, теплицы) можно регулировать также воздушное П. р.— путём изменения содержания CO2 в воздухе и дополнительного освещения (см. Светокультура растений). Создание оптимального комплекса условий для П. р.— главная задача растениеводства. На решение этой задачи направлены мероприятия по мелиорации засоленных почв (удаление вредного для П. р. избытка солей), агротехнические приёмы обработки почвы (создание условий плотности и аэрации, облегчающих П. р.), борьба с сорняками (конкурирующими с культурными растениями за элементы П. р.) и др.
Лит.: Тимирязев К. А., Жизнь растений, Избр. соч., т. 3, М., 1949; Сабинин Д. А.. Физиологические основы питания растений, М., 1965; Максимов Н. А., Как живёт растение, 4 изд., [М., 1966].
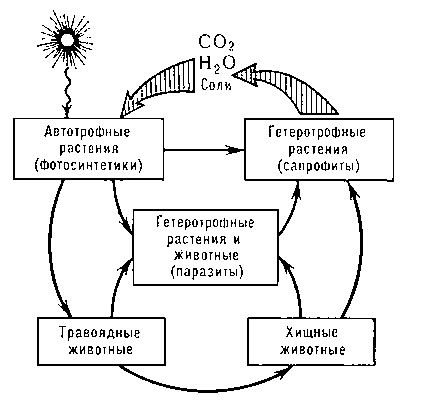
Рис. 1. Биогеохимический круговорот веществ в природе.
Фотосинтез – Превращение и передвижение ассимилятов у винограда
Содержание материала
Синтезированные ассимиляты не задерживаются в листьях, а передвигаются к точкам роста и к ягодам, где используются для ростовых процессов и формирования урожая. Во время вегетации и, особенно, после прекращения роста продукты фотосинтеза в форме полисахаридов (крахмал, гемицеллюлоза, лигнин и целлюлоза) отлагаются в больших количествах в корневой системе, в многолетней древесине и однолетнем приросте. В осенне-зимний период крахмал, находящийся в надземных частях, превращается в сахара, выполняющие защитные функции, а в корневой системе сохраняется в неизменившемся состоянии как резервное вещество. В начале вегетации крахмал, накопленный в корнях, начинает перемещаться к развивающимся почкам и молодым побегам. Передвижение ассимилятов происходит и в многолетних частях, и в однолетних побегах.
Передвижение ассимилятов в восходящем и нисходящем направлении, как и их транслокация в отдельных органах и частях, сопровождается непрерывным превращением углеводов. Поэтому изучение путей передвижения ассимилятов, как и их превращения, представляют значительный теоретический и практический интерес, особенно с целью регулирования количества и качества урожая.
Природа продуктов фотосинтеза и органических веществ, переносящих ассимиляты. Природу продуктов ассимиляции винограда изучали немногие исследователи. М. Венециа (Venezia, 1938), определяя содержание сахаров (глюкозы, фруктозы и сахарозы) в листьях различных сортов винограда в ранние утренние и послеполуденные часы, констатирует значительное увеличение только сахарозы. На основании этих результатов он приходит к заключению, что сахароза накапливается в солнечные часы и что между содержанием ее и фотосинтетической деятельностью листьев существует тесная связь. Автор даже делает вывод, что сахароза предшествует всем другим сахарам и первая образуется в листьях во время фотосинтеза. Вывод Венециа об определенной связи между содержанием сахарозы и фотосинтетической активностью листьев не утратил научного значения и в настоящее время (Calvin, 1962). В исследованиях М. В. Туркиной (1959), например, доказывается, что радиоактивный углерод накапливается главным образом в сахарозе и совсем в незначительных количествах в глюкозе и фруктозе.
Некоторое время после подкормки радиоактивным углеродом содержание его в глюкозе и фруктозе увеличивается. Это, однако, наступает после расщепления сахарозы. Поэтому в настоящее время сахароза считается одним из первых продуктов фотосинтеза.
Она является важнейшим растворимым сахаром, содержащимся в листьях (Marteau, 1960). Днем ее больше, чем в конце ночи. Из этого следует, что в течение светового периода в листьях накапливается запас сахарозы, который исчерпывается ночью. Отношение, установленное в листьях в 14 ч (1,78), всегда выше, чем в 4 ч утра (1,18). К концу послеполуденного периода сахароза больше чем вдвое превышает количество редуцирующих сахаров, в то время как рано утром их почти одинаковое количество.
По схеме Кальвина (Calvin, 1962), из первого продукта фотосинтеза — фосфоглицериновой кислоты путем дальнейшего трансформирования последовательно образуются фосфоглицеринальдегид и фосфодиоксиацетон. Под действием альдолазы из них образуется фруктозо-1,6-дифосфат, из которого синтезируются сахароза и различные полисахариды.
Известные данные о характере ассимилятов в листьях винограда получены Клеммом (Klemm, 1958). После подкормки листьев 7—9-недельных привитых саженцев в течение 3,5 ч С1402 он с помощью методов радиохроматографии разделил и идентифицировал вещества, имеющиеся в листьях и содержащие радиоактивный углерод. Среди идентифицированных веществ установлено наличие яблочной, лимонной, винной и глюкуроновой кислот, глюкозы и фруктозы. Количественное соотношение идентифицированных веществ, следующее (в % общей хроматографическоп активности):
глюкуроновая кислота и неидентифицированные вещества
с почти одинаковыми значениями. 14±5
другие неидентифицированные вещества. 11+3
глюкоза. 28+4
фруктоза . 18+4
винная кислота. 5,5+2
лимонная кислота. 10+2
яблочная кислота. 14+4
Соотношение сахаров и кислот составляет приблизительно 3 : 2. Радиоактивный углерод за время экспозиции успел войти в состав клетчатки листьев.
Исследования Клемма не доказывают возможность образования различных продуктов фотосинтеза, так как экспозиция длилась 3,5 ч. По данным А. А. Ничипоровича (1955), М. Кальвина и Д. Басхэма (1956), вместе с продуктами фотосинтеза углеводного типа в процессе ассимиляции наибольшее количество С14 включается в аминокислоты — аланин, серин, аспарагиновую и глютаминовую кислоты. Однако интенсивное увеличение количества радиоактивного углерода в составе этих аминокислот наблюдается лишь после 5-минутной экспозиции, т. е. когда все промежуточные продукты фотосинтеза были насыщены С14.
При изучении продуктов фотосинтеза 27 растений, листья которых были экспонированы в среде радиоактивной углекислоты в течение 10—60 сек, определены гексозомонофосфаты, пентозы и триозофосфаты, уридин-глицериновая, аспарагиновая, глютаминовая кислота, серии, аланин, моно- и диса-хариды и органические кислоты (L. Noris, Е. Noris, 1955).
Значительно полнее природу ассимилятов, образованных в листьях винограда, выяснил Ш. Ш. Чанишвили (1964). Он установил, что после 5-минутной экспозиции листьев сорта Ркацители в атмосфере с С1402 87,4% радиоактивного углерода включается в углеводы, 7,8% в органические кислоты и 2,1 % в аминокислоты. Основная доля радиоактивного углерода сохраняется в углеводах и после более продолжительной экспозиции: 30 мин — 81,9%; 60 мин — 76,3%; 3 дня — 59,9%. С увеличением экспозиции повышается доля радиоактивного углерода в органических кислотах: 30 мин— 10,7%; “60 мин— 11,4% и 3 дня — 18,7%- Увеличивается также его количество в аминокислотах при экспозиции в течение 3 дней до 7%.
Основная часть радиоактивного углерода, ассимилированного листьями, включается в состав сахарозы. Содержание С14, включенного в глюкозу, фруктозу и рафинозу, примерно в 2—2,5 раза меньше.
Кроме продуктов углеводного типа Ш. Ш. Чанишвили установил в листьях винограда целый набор аминокислот и органических кислот, содержащих радиоактивный углерод.
Из аминокислот наиболее радиоактивными оказались аспарагиновая кислота, пролин и тирозин, а из органических — винная и яблочная (табл. 76).
Фиксирование С02 в течение 3—30 сек было проведено Риберо-Гайоном (Ribereau-Gayon, 1966). После 10-секундной экспозиции меченый углерод (С14) обнаруживается в сложных фосфорных эфирах седогептулозы, глюкозы и фруктозы в форме монофосфатов (41%) и в фосфороглицериновой кислоте (37%).
При фиксировании С02 в течение более продолжительного времени (через 6, 30 мин или 2 ч) наибольшая доля радиоактивности содержится в углеводах. За 30 мин сахароза содержит уже 64% всей радиоактивности углеводов, фруктоза—15% и глюкоза — 13%. При 2-часовой экспозиции относительное содержание сахарозы во взрослых листьях уменьшается (11%), а в молодых составляет 59%. Так как содержание сахарозы колеблется в течение суток, некоторые авторы считают сахарозу важнейшим сахаром в листьях винограда (Guichard, 1954; Marteau, 1955).
Существенным является вопрос о природе веществ, переносящих ассимиляты.
Исследования К. Соунсона (1961) показали, что содержание сахаров (меченых+немеченых) изменяется по длине стебля, причем количество сахарозы над местом подкормки С1402 медленно увеличивается, а глюкозы и фруктозы уменьшается. В коре побега в значительной степени преобладает сахароза, а в ксилеме — глюкоза (табл. 77).
Таблица 76
Набор органических кислот и аминокислот в листьях и распределение в них С14 
Таблица 77
Зависимость концентрации сахаров (меченых и немеченых) в стебле от расстояния до места подкормки См02
Источники:
http://naukarus.com/osobennosti-sutochnogo-ritma-peredvizheniya-assimilyatov-v-vinogradnoy-loze
http://gufo.me/dict/bse/%D0%9F%D0%B8%D1%82%D0%B0%D0%BD%D0%B8%D0%B5_%D1%80%D0%B0%D1%81%D1%82%D0%B5%D0%BD%D0%B8%D0%B9
http://vinograd.info/info/fiziologiya-vinograda/fotosintez-2.html





