
Метод определения продуктивности фотосинтеза листьев винограда в связи с урожаем
Метод определения продуктивности фотосинтеза листьев винограда в связи с урожаем
Фотосинтетическая деятельность и продуктивность виноградника – Вопросы методики учета фотосинтетической деятельности винограда
Содержание материала
IV.1. НЕКОТОРЫЕ ВОПРОСЫ МЕТОДИКИ УЧЕТА ФОТОСИНТЕТИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ВИНОГРАДА В НАСАЖДЕНИЯХ
В настоящее время разработаны общие принципы и методы учета ФД растений в посевах [201, 260, 293]. Применительно к винограду параметры некоторых из них требуют уточнения, в частности определение площади листьев, ЧПФ и др.
Существует большое число методов определения площади листьев с учетом их морфологии [260, 306, 383]. В исследованиях с виноградом в зависимости от требуемой точности и условий работы целесообразно использовать следующие методы: планиметрирования, весовой, а также различные модификации расчетных методов, в основе которых лежит корреляционная связь между площадью листа и некоторыми его морфологическими параметрами: диаметром, длиной боковых жилок и др. [186, 199, 421, 464].
В основе широко распространенного в виноградарстве «ампелометрического» метода [199] лежит представление о площади листа как площади круга. Поскольку фактическая площадь листа меньше площади круга, вносится поправка на фирму листа и его рассеченность. Поправочный коэффициент рассчитывается как отношение величины истинной площади липа, полученной с помощью планиметра, к вычисленной по формуле. В условиях Крыма для группы сортов поправочный коэффициент изменяется от 0,82 до 0,88 [15], для группы сортов и Грузии [37] — от 0,74 до 0,79. Достоинством расчетного метода является простота, достаточно высокая точность и возможностиопределения площади без отделения листа от растения.
Площадь листьев побега определяется суммированием площадей всех листьев побега.
Эффективны также расчетные методы, основанные ми линейной зависимости между длиной побега и площадью его листьев [173]. На рис. 24 показана такая зависимость для ряда сортов винограда в условиях Крыма. В конце вегетации в зависимости от погодных условий количество листьев на побегах может варьировать, потому целесообразнее рассчитывать площадь листьев побега умножением количества листьев на площадь «среднего» листа. Площадь листьев всего куста получают умножением площади листьев «среднего побега» на среднюю нагрузку побегами.
Для характеристики ФД растений наряду с размерами площади листьев используется показатель фотосинтетического потенциала (ФП). Он выражается в м 2 >днях и представляет собой сумму ежедневных площадей листьев растения или ценоза за вегетационный период или за какую-то его часть. Величина ФП побега, куста или виноградника рассчитывается суммарно за вегетацию или за отдельные его периоды, как это показано на рис. 25. Площадь фигуры АБВ определяют с помощью планиметра или весовым методом. Общим требованием при проведении биометрических измерений является представительность выборки [379].
У винограда подавляющая часть ассимилятов используется пи построение побегов и гроздей. Часть ассимилятов затрачивается на рост корней, утолщение штамба, рукавов и скелетных корней, а также откладывается в них в форме запасных веществ. Определение всей массы годичной фотосинтетической продукции в многолетних частях куста связано с большими методическими и техническими трудностями. Поэтому в исследованиях за величину биологической продукции Убиол обычно принимают массу одногодичных побегов (с листьями) и гроздей. Хозяйственно ценную часть урожая УХоз составляют грозди.

Рис. 24. Зависимость площади листьев, побега от его длины i.Сорта: 1 — Хиндогны, 2 — Мускат белый (Южный берег Крыма), 3 — Шабаш, 4 — Рислинг рейнский, 5 — Ркацители (западная степная прибрежная зона Крыма).

Рис. 25. Пример определения величины фотосинтетического потенциала ФП по площади фигуры АБВ (по [143]).
В исследованиях с виноградом получили распространение методы определения фотосинтетической активности листьев, используемые обычно в работе с другими культурами [78, 260]. Это главным образом различные модификации газометрического и весового методов, определение фотосинтеза с помощью
14СС>2 и др. Однако, изучение взаимосвязи фотосинтеза с урожайностью не может ограничиваться только учетом количества усвоенной СО2 [250]. Более того, прямые опыты показали [320], что между уровнем наблюдаемого фотосинтеза и накоплением биомассы винограда нет прямой коррелятивной связи. Это особенно относится к кратковременным и единичным измерениям и к случаям, когда сравниваются сорта с различными биологическими свойствами.
Причина отсутствия такой связи — прежде всего чисто методическая. Правомерно в связи с этим замечание Стоева [336] о том, что путем определения фотосинтеза отдельного листа можно получить сведения о метамерных, сезонных и суточных изменениях фотосинтеза листьев. Однако эти данные не могут создать представления о суммарном фотосинтезе всего куста за суточный цикл и балансе распределения и использования ассимилятов на формирование гроздей и вегетативных органов. И совершенно очевидно, что такие кратковременные определения фотосинтеза на отдельных листьях не могут служить для обоснования каких-либо выводов, касающихся величины урожая и его качества, как это иногда еще имеет место в ряде работ (большей частью агротехнического характера).
В исследованиях по проблеме «Фотосинтез и урожай» для характеристики фотосинтетической активности листьев в связи с накоплением биомассы растения чаще всего используется показатель ЧПФ, характеризующий весовое количество суточного прироста сухого вещества растения в расчете на 1 м 2 площади листьев. Величина ЧПФ может рассчитываться как средняя за вегетацию, так и за отдельные его периоды на общую биомассу или на хозяйственную часть (ЧПФХ03), характеризуя тем самым роботу листового аппарата на формирование гроздей.
Вместе с тем следует отметить, что величина ЧПФ — не однозначный показатель фотосинтетической активности листьев. Она может меняться при постоянном фотосинтезе в зависимости от дыхания, а также тогда, когда фотосинтез и дыхание стабильны, но изменяется отношение сухой фитомассы растений к его листовой поверхности; при густом РП общая площадь листьев может быть больше фактической ассимилирующей листовой поверхности; если фотосинтезируют и стебли растений, то величина ЧПФ, рассчитанная на единицу площади листовой поверхности, будет завышена. Эти особенности пока ЧПФ следует принимать во внимание при интерпретации получаемых данных.
В исследованиях с виноградом для характеристики работы листьев в связи с урожаем гроздей часто используется показатель продуктивности листьев (ПЛ). Величина ПЛ рассчитывается как отношение сырой массы гроздей или сахара гроздей к единице площади листьев куста
Следует, однако, отметить, что простой пересчет сырой массы гроздей (или сахара гроздей) на площадь листовой поверхности отражает не потенциальную продуктивность единицы площади листьев, а фактически сложившееся соотношение между массой гроздей и листовой поверхностью. Иначе говоря, такой пересчет дает представление о степени использования листовой поверхности на формирование гроздей, но не характеризует работу единицы площади листьев как таковой. Очевидно, что при недогрузке куста гроздями образуется «избыток» площади листьев и величина ПЛ окажется заниженной. При перегрузке гроздями в формирование их частично вовлекаются пластические вещества из многолетних частей куста [49, 238] и величина ИЛ завышается. Методика определения потенциальной ПЛ на изолированных кольцеванием побегах [15] учитывает эту особенность винограда.
Интегрирующим показателем ФД растений в фитоценозах является коэффициент использования падающей ФАР щ. Он характеризует эффективность использования ценозом падающей ФАР на всю площадь РП в зависимости от видового состава, структуры РП и др.
1 Поскольку процентное содержание сахара в соке ягод в разных случаях разнос, правильнее характеризовать этот показатель не по сырой массе, и по количеству сахара гроздей.
КПД ФАР служит мерилом относительного уровня использования солнечной энергии фитоценозом, но не дает исчерпывающей характеристики продуктивности ценоза как источника того продукта, ради которого культура возделывается. Поэтому целесообразно одновременно иметь данные о величине и качестве хозяйственной продукции.
В настоящее время разработана система объективной оценки фотосинтетической продуктивности ценозов [251, 255]. Продуктивность, при которой растения посева или насаждения накапливают приходящую за время фактической вегетации ФАР с суммарным КПД всего 0,5—1%, считается низкой; продуктивность посевов с КПД 1—2%—средней, 2—3%—хорошей, 3—4%—высокой и 4—5%—очень высокой. В исследованиях, проведенных в контролируемых условиях при оптимизации всех факторов среды (где лишь ФАР являлась лимитирующим фактором), показано, что КПД ФАР по продуктивности ценоза в широком диапазоне ее интенсивности (
50—150 вт/м 2 ) составил примерно 6%, что отвечает теоретически максимальной величине [226]. В качестве возможных принимаются следующие КПД падающей ФАР на формирование урожаев: в зонах полной обеспеченности влагой, а также в культуре растений с поливами — не менее 4 и до 5—6%; в зонах недостаточного увлажнения (средняя зона) — 1—2%; в сухих зонах (сухая степь, полупустыня) —0,7—1,0% [251, 255].
Как известно, калорийность органических соединений определяется их составом. Растения, содержащие повышенное количество жира и белка, имеют высокую калорийность (5,5—6,5 ккал/г), углеводистые — относительно низкую (4,0— 4,3 ккал/г), зерновые культуры по теплотворности занимают промежуточное положение — 4,5—5,5 ккал/г [160].
Таблица 14
Калорийность органов винограда1 в момент уборки урожая (ккал/г сухого вещества)
Метод определения продуктивности фотосинтеза листьев в связи с урожаем
Существующие методы определения продуктивности фотосинтеза листьев у растений можно разделить на следующие группы.
- Газометрический метод со всеми его модификациями. Для определения фотосинтеза воздух или газовую смесь пропускают через камеру с листом или растением и определяют концентрацию СО2 во входящем и выходящем из камеры воздухе.
- Определение фотосинтеза по изменению радиоактивности газовой смеси, содержащей C14O2, которую пропускают через камеру с листом или растением.
- Определение количества выделяющегося при фотосинтезе кислорода.
- Различные модификации определения фотосинтеза по накоплению ассимилятов в листьях растения.
Каждый из этих методов в большей или меньшей степени применяют в различного рода физиологических исследованиях как в стационарных, так и в полевых условиях.
Сравнительно новое направление в изучении фотосинтеза, выросшее в проблему фотосинтеза и урожая, ставит своей задачей выявление количественной связи между фотосинтезом и накоплением всей массы или хозяйственно ценной части растения. Фотосинтез выступает здесь не изолированно, а в тесной взаимосвязи с
ростом и формированием листового аппарата растения, с продолжительностью работы листьев на создание урожая, с величиной и качеством получаемой продукции, то есть как составной элемент интегрального процесса формирования урожая.
Разработка проблемы фотосинтеза и урожая требует новых подходов, новых методов учета фотосинтетической деятельности листьев растений в посевах и насаждениях, поскольку существующие методы в данном случае оказываются малоэффективными. Следующие обстоятельства объясняют это положение.
- Величина фотосинтеза, полученная на единичных листьях, характеризует работу самого листа за короткий период (обычно за несколько часов) безотносительно к накоплению органической массы всего растения и не может дать представления о действительной работе листьев в течение всего дня и тем более всего периода вегетации. В случае пересчета на площадь листьев всего растения допускаются существенные ошибки
- Названные методы дают величину наблюдаемого или кажущегося фотосинтеза. Для установления истинного фотосинтеза необходимо вводить поправку на дыхание и отток, происходящие одновременно с фотосинтезом на свету.
- Указанные методы не учитывают качественную сторону работы листьев, выражением которой является коэффициент эффективности фотосинтеза – Кэф, (А.А. Ничипорович и др., 1938) _ отношение количества сухой массы урожая к весовому количеству усвоенной СО2. Коэффициент этот сильно варьирует (от отрицательных величин до +60%) в зависимости от направленности работы листьев, характера утилизации продуктов фотосинтеза, дыхания и т.п.
- Между интенсивностью фотосинтеза и накоплением органической массы растения нет прямой связи. Урожай, помимо фотосинтеза, зависит от величины, характера роста и формирования листового аппарата, продолжительности работы листьев на формирование урожая. Величина хозяйственно ценной части урожая (зерно, плоды) зависит еще от структуры растения – соотношения между урожаем плодов и общей массой биологического урожая, или иначе – коэффициента хозяйственной эффективности фотосинтеза – К хоз.
В исследованиях по проблеме фотосинтеза и урожая широкое применение получил метод учета чистой продуктивности фотосинтеза, характеризующий чистый прирост сухого вещества всего растения в расчете на среднюю площадь листьев за определенный отрезок времени, обычно 7-10 дней (А.П.Бегишев, 1853; А.А. Ничипорович, 1961). В связи с изменением площади листьев растения происходящим за вегетацию, значительный интерес представляет опенка фотосинтетической деятельности листьев по величине фотосинтетического потенциала. Продуктивность фотосинтеза в этом случае выражается в м2 -днях, необходимых для получения единицы сухой массы растения.
В основу разработанного и предлагаемого нами метода положены два последних принципа учета фото— синтетической деятельности листьев. Продуктивность их работы у винограда в связи с урожаем гроздей характеризуют по фотосинтетическому потенциалу, необходимому для получения 1 кг сахара (ФП в м2-днях), и по средней величине чистой продуктивности фотосинтеза за период формирования урожая (Ф в г сахара/м2-день).
За период формирования урожая у винограда условно принимается время от момента завязывания ягод До наступления полной их зрелости, когда абсолютное накопление сахара в ягодах прекращается. Наблюдения за сахаронакоплением проводят на нормально развивающихся плодоносных побегах, которые берут в качестве контрольных.
Продуктивность фотосинтеза листьев винограда в связи с урожаем можно в принципе определять и на целых кустах. Однако для этого необходимо, чтобы было установлено и поддерживалось оптимальное соотношение между площадью листьев и урожаем гроздей куста , что на практике трудно выполнимо. В случае недогрузки урожаем окажется некоторый избыток площади листьев и показатели продуктивности будут занижены.
При перегрузке показатели окажутся завышенными, так как в формирование урожая частично будут вовлечены органические вещества из многолетних частей куста и корней. Кроме того, работа с целыми кустами сама по себе затруднительна вследствие громоздкость и трудоемкости учетов.
Сущность предлагаемого метода состоит в изоляции путем кольцевания группы однолетних побегов от остальной части куста и учета произведенной листьями этих побегов продукции по количеству валового сахара формирующихся на них гроздей.
Сразу же после цветения у основания плодового побега снимают кольцо луба шириной около 0,5 см. Что бы предупредить подсыхание древесины и возможность срастания каллюса, место кольцевания обматывают изоляционной лентой или медицинским пластырем. В период вегетации, в случае необходимости, вырез кольца обновляют или делают новое кольцо, а старому дают срастись. На плодовом побеге оставляют 3-4 плодоносных побега с одинаковым числом листьев и с 1 гроздью на каждом побеге. Листья равномерно распределяют по всей длине побега. Обязательным является условие, чтобы площадь листьев ограничивала рост урожая. Как показали наши исследования, именно в этих пределах величина продуктивности сохраняется примерно одинаковой. В зависимости от величины грозди 6-8 Листьев на побеге ограничивают рост урожая. Проявляется это в уменьшении размера опытной грозди и ягод в ней по сравнению с контрольными. Верхушку побега
и пасынки удаляют. В результате вся продукция, вырабатываемая листьями побега, поступает в гроздь, а также частично расходуется на утолщение стебля и листьев. Работа листьев оценивается по валовому содержанию моносахаров в соке ягод и легкогидролизуемых углеводов в семенах и кожице с плотными частями мякоти, также выраженных в моносахарах. Углеводы, имеющиеся в гребнях, можно не учитывать, так как они составляют очень небольшую часть от массы грозди. В некоторых случаях для упрощения валовой сахар можно определять только по содержанию его в соке тщательно отжатых ягод. Площадь листьев определяют в момент завязывания ягод, 2-3 раза в период формирования урожая и при съеме урожая. Все учеты и определения проводят по каждому плодоносному побегу в отдельности. Таким образом, побег с гроздью является учетной единицей, повторностью.
Д я получения показателей продуктивности фотосинтеза строят график, на оси абсцисс которого наносят в масштабе даты начала формирования и съема урожая, а на оси ординат – площадь листьев побега в соответствующие даты измерения (рис.). Концы отрезков, соответствующие площадям листьев, соединяют друг с другом и получают замкнутую площадь, например АВСД, Величину ФП получают путем умножения площади АВСД на показатели цены 1 см масштаба на оси абсцисс и на оси ординат. Например:
площадь фигуры АВСД-23,7 см ,
1 см на оси абсцисс – 10 дням,
1 см на оси ординат -2 дм,
ФП побега – 23,7х 10х2-474 дм2-дней, или
4,74 м2-дней (АX. Амирджанов, 1963).

Расчет величины фотосинтетического потенциала по площади фигуры АВСД.
Это значит, что гроздь с валовым сахаром 9,68 г была создана в результате работы листьев, равной 4.74 -дней. По этим данным рассчитывают величину ФП, необходимую для получения 1 кг сахара. В рассматриваемом случае для получения 1 кг сахара необходим ФП, равный 480 м-дней.

Второй показатель продуктивности, среднюю за период формирования урожая величину чистой продуктивности фотосинтеза, получаем делением валового сахара грозди на ФП побега.
Полученные этим методом показатели продуктивности фотосинтеза для сортов винограда в условиях Степного Крыма и Южного берега Крыма даны в таблице 1.
Метод кольцевания может, на наш взгляд, дать вполне удовлетворительные результаты при изучении таких вопросов, как продуктивность фотосинтеза у разных сортов винограда, у листьев разных ярусов, у листьев одного и того же яруса на протяжении вегетации при воздействии на растения стимуляторами, микроэлементами и др. Действительно, в этих случаях определенная площадь листьев производит во времени конкретную массу урожая. И то, в другое поддается точному количественному учету и объективному выражению.
Таблица 1
Продуктивность фотосинтеза листьев винограда

Имеется и другая большая категория исследований, где могут быть использованы показатели продуктивности фотосинтеза. Сюда относятся работы, связанные с зелеными частями растения: определение оптимальной нагрузки кустов побегами и гроздями, высоты и сроков прищипки и чеканки побегов и т.п. В этих работах приходится оперировать целыми растениями и, естественно, возникает вопрос: в какой мере показатели продуктивности фотосинтеза, полученные методом кольцевания, могут быть перенесены на целый куст? В таблице 2 приведены данные по продуктивности фотосинтеза, полученные путем учета прироста всей надземной части виноградного растения в расчете на единицу площади листьев и методом кольцевания.
Таблица 2
Продуктивность фотосинтеза листьев винограда (г сухого вещества/ м2 -день)
Методика определения чистой продуктивности фотосинтеза растений.
Отбирают три растения с каждого варианта условий. Все зеленые листья отрывают от растения, взвешивают и складывают в бюксы. Затем взвешивают все стебли и складывают в другие бюксы.
Далее определяют площадь зеленых листьев. После этого бюксы с растительной массой ставят в сушильный шкаф и сушат до воздушно-сухого состояния (105 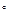 С). Затем растительный материал взвешивают, определяя воздушно-сухую массу зеленых листьев и стеблей.
С). Затем растительный материал взвешивают, определяя воздушно-сухую массу зеленых листьев и стеблей.
Через 3-4 дня вновь берут такое же количество растений с каждого варианта условии. Растения также разбирают, раскладывают в бюксы, определяют их сырую массу, площадь зеленых листьев, ставят в сушильный шкаф, просушивают и взвешивают.
Чистую продуктивность фотосинтеза растений по формуле Веста- Брикса.
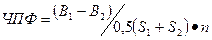 , где
, где
ЧПФ – чистая продуктивность фотосинтеза, г/м 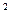 в сут.,
в сут.,
В 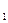 , В
, В 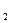 – сухая масса растений в конце и в начале учетного периода, г.,
– сухая масса растений в конце и в начале учетного периода, г.,
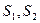 – площадь зеленых листьев в начале и в конце учетного периода, м ,
– площадь зеленых листьев в начале и в конце учетного периода, м ,
n – число дней между двумя определениями [Викторов].
Во время летней практики было проведено ряд измерений для описаний различных видов и сортов злаковых растений, на примере «Пшеница яровая (контроль)» и «Пшеница яровая тулейковская (опыт)». Мы брали три пробы в разное время лета: 30.06, 08.07 и 15.07. Каждый раз мы измеряли 3 растения для более средней и точной характеристики сорта.
1) мы измеряли площадь каждого листа, для определений средней площади, определяли их массу и влажность, для расчеты фотосинтетической продуктивности растений (методику см.выше).
2) проводились измерения корневой системы. Измерялась ее длина, масса и объем. Объем измерялась по следующей схеме. Определение объема корневой системы методом Д.А. Сабинина и И.И.Колосова. (методику см. ниже). Также готовились мазки из смывов с корней, их окраска по Граму.
3) проводились исследования колоса растений, а именно массы, количества зерен и определение влажности.
Первый отбор – 30.06.2011.
1. Пшеница яровая (контроль):
Растение №1. Площадь листьев: S =0.7•25.5•0.9=16
S =0.6•19•0.9=5.1 S 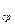 =8.4м
=8.4м
S 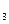 = 0.2•23•0.9=4.1
= 0.2•23•0.9=4.1
Источники:
http://vinograd.info/info/solnechnaya-radiaciya-i-produktivnost-vinogradnika/fotosinteticheskaya-deyatelnost-i-produktivnost-vinogradnika-2.html
http://vinograd.info/knigi/metodiki-po-fiziologo-biohimicheskim-issledovaniyam/metod-opredeleniya-produktivnosti-fotosinteza-listev-v-svyazi-s-urozhaem.html
http://mylektsii.ru/2-95023.html





