
Краткие сведения об анатомическом строении семян – Физиология роста и развития семян и сеянцев винограда
Краткие сведения об анатомическом строении семян – Физиология роста и развития семян и сеянцев винограда
Физиология размножения винограда семенами
Содержание материала
КРАТКИЕ СВЕДЕНИЯ ОБ АНАТОМИЧЕСКОМ СТРОЕНИИ СЕМЯН
Семена винограда относятся к анатропным. Они овально-грушевидной формы с удлиненным клювиком. Внешний вид и величина семян видов и сортов рода Vitis являются довольно постоянными ампелографическими признаками. Семя V. vinifera имеет длинный клювик и халазу, расположенные в верхней трети спинной стороны. Клювик семян других видов этого рода более короткий, а халаза находится в средней части спинной стороны.
Семя винограда состоит из внешнего и внутреннего покровов, эндосперма и зародыша. Внешний покров образуется из внешнего интегумента семяпочки. В нем различают внешний, промежуточный и внутренний слои. Внешний слой состоит из одного ряда тангентально расположенных клеток, которые содержат крахмал и рафиды. Клетки среднего слоя тонкостенные, рыхлого строения. У несозревших семян они содержат в большом количестве крахмал и рафиды, а у созревших — высыхают, сплющиваются и становятся вместилищем танина. Этот слой легко поглощает воду и увеличивает свою толщину, поэтому его называют губчатым. Внутренний слой образуется из внутреннего эпидермиса и состоит из 2—6 слоев радиально удлиненных каменистых клеток. Оболочка из каменистых клеток почти полностью окружает семена винограда, свободными от нее остаются только халаза и верхушка клювика, где находится отверстие, называемое микропиле. В различных зонах семени число каменистых клеток неодинаково: в клювике около микропиле их больше, а в средней части — меньше.
Внутренний покров образуется из внутреннего интегумента семяпочки и также состоит из трех слоев, каждый из которых имеет один слой клеток. Внешний слой (эндодермис) представляет собой сильно сплющенные тонкостенные клетки, а средний — тонкостенные многоугольные клетки. Внутренний слой плотно прилегает к эндосперму. Клетки этого слоя подвержены значительным изменениям. При созревании семян на радиальных стенках их появляются утолщения, как и в каменистых клетках. В полностью созревших семенах стенки этих клеток бурые, поэтому эндосперм резко отграничен.
Эндосперм занимает внутреннюю часть семян. Он состоит из множества крупных клеток с утолщенными в углах стенками. Клетки содержат в избытке капли жира, белковые вещества, протоплазму и минеральные соли.
Зародыш находится в клювике, корешок обращен к микропиле. Две семядоли зародыша имеют удлиненную сердцевидную форму. Между ними находится небольшая зародышевая почечка — эпикотиль (столбик), цилиндрической формы, заканчивающийся корешком.
ХИМИЧЕСКИЙ СОСТАВ СЕМЯН И ОБМЕН ВЕЩЕСТВ В НИХ
По данным О. П. Смульской (1961), в химическом составе семян разных сортов существенных различий нет (табл. 57).
Большое внимание ряд авторов уделяли изучению содержания дубильных веществ в семенах винограда. Н. М. Сисакян, И. А. Егоров и Б. Л. Африкян (1947, 1948) установили, что нарастание количества дубильных веществ в семенах винограда до начала созревания совпадает с уменьшением их содержания в листьях, гребнях и ягодах винограда.
Таблица 57 Химический состав семян разных сортов винограда (в % на сухое вещество) 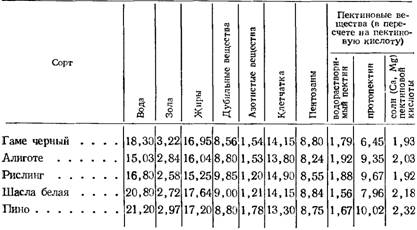
Чтобы выяснить, не вызвано ли нарастание дубильных веществ в семенах оттоком их из мякоти, кожицы и гребней, С. В. Дурмишидзе и М. П. Цискаришвили (1950) изучали количественные изменения этих веществ в бессемянных сортах. Полученные ими результаты показали, что уменьшение содержания дубильных веществ в мякоти, кожице и гребнях нельзя объяснить оттоком танидов в семена.
Снижение содержания дубильных веществ в семенах винограда после начала созревания ягод обнаружено Н. М. Си-сакяном (1947), С. В. Дурмишидзе (1950), К. А. Стоевым и И. Димитровым (1957) и др. По мнению Н. М. Сисакяна, оно указывает на вовлечение этих соединений в обмен веществ и на их отток в одревесневшие части растения. По данным С. В. Дурмишидзе и Т. П. Цискаришвили (1950), в семенах винограда в стадии технической спелости остается в среднем 40—60% первоначального количества дубильных веществ, причем соотношение между водорастворимыми и щелочера-створимыми танидами в одних сортах (Ркацители, Мцване, Чинури, Горула, Тавквери-картлури, Шавкапито, Симона-сеули, Горула-Мцване и др.) возрастает, а в других (Буде-шура-тетри,” Цулукидзис-тетри, Кумси-тетри, Каси и Шаба) почти не изменяется.
Процесс снижения количества дубильных веществ в семенах продолжается и после снятия гроздей с куста (Дурмишидзе, Цискаришвили, 1960). После 5-месячного хранения гроздей содержание водорастворимых танидов в семенах снизилось на 19% (табл. 58).
Таблица 58
Содержание дубильных веществ в семенах сорта Ркацители при хранении винограда (в мг на I г сухого вещества)
Физиология роста и развития семян и сеянцев винограда – Особенности роста и развития сеянцев винограда в зависимости от длины дня
Содержание материала
ОСОБЕННОСТИ РОСТА И РАЗВИТИЯ СЕЯНЦЕВ В ЗАВИСИМОСТИ ОТ ДЛИНЫ ДНЯ (ФОТОПЕРИОДИЗМ)
В своем индивидуальном развитии сеянцы винограда, так же, как и остальных многолетних древесных растений, предъявляют определенные требования к продолжительности солнечного освещения (к длине дня).
Рассматривая вопрос об историческом развитии винограда, Давитая (1948) приходит к заключению, что в более ранние геологические периоды виноград занимал значительно более широкие географические ареалы, чем сейчас. Поэтому он считает, что фотопериодическая амплитуда винограда должна быть значительно более широкой по сравнению с ее пластичностью в отношении температурного фактора.
Исследования, посвященные фотопериодизму винограда, немногочисленны. Первые исследования в этом направлении — это опыты Хакбарта и Шерца (цит. по Мержаниану, 1939) и Потапенко и Захаровой (1937). Ими было установлено, что длина дня оказывает существенное влияние на рост и развитие сеянцев винограда. При более коротком дне сеянцы как будто приобретают более короткий вегетационный период и более высокую морозоустойчивость. Кроме того, их корневая система развивается сильнее.
Позже Соловей (1958) установил, что т. наз. световая стадия у сеянцев винограда проходит при естественной длине дня (15—16 h) после воздействия света на протяжении 40—50 дней. Сеянцы, выращиваемые в условиях короткого дня (8—12 h), до конца вегетационного периода не приходят в готовность к плодоношению в следующем году.
Гречишников (1957), изучавший влияние продолжительности дня на вызревание побегов, устанавливает, что в условиях короткого дня побеги начинают вызревать значительно раньше, чем при нормальной продолжительности дня. При этом степень фотопериодического воздействия зависит от его продолжительности. Фотопериодическое воздействие на протяжении одного-двух дней не приводит к ускорению вызревания побегов, 3—4-дневное вызывает ускорение вызревания на 5—12 дней, 5—6-дневное — на 16—21 день, а 7-дневное и больше — на 30 дней.
Гречишников установил также, что накопление углеводов, и в особенности крахмала, протекает интенсивнее у сеянцев, выращиваемых в условиях короткого дня, в результате чего их морозостойкость повышается. Укорачивание дня, однако, приводит и к значительному замедлению роста побегов. Аналогичная констатация сделана также Аллевельдтом (1960). Давитая (1948) в свою очередь считает, что разница в длине дня до 3 h (при продолжительности дня 12—15 h) не оказывает существенного влияния на урожай и сроки созревания винограда.
В соответствии с симподиальной теорией рост побегов винограда V. vinifera происходит при непрерывном чередовании междоузлий с усиками с междоузлиями без усиков. Сначала рост побегов осуществляется моноподиально, т.е. от верхушечной меристемной почки вверх. После образования 3—5-го междоузлий, однако, почка роста конуса нарастания сдвигается в сторону, преобразуясь в усик, а рост осуществляется от новой почки роста, возникшей на обратной стороне. В следующем междоузлии, однако, и эта почка сдвигается и переходит в усик, а рост кверху осуществляется от почки роста, возникшей также на обратной стороне. Эта почка осуществляет рост побегов двух последующих междоузлий моноподиально, в результате чего находящийся между ними узел не имеет усика. Дальше описанный цикл чередования моноподиального роста с симподиальным повторяется в строгой последовательности, в результате чего на нормально развитом побеге через каждые два супротивно расположенные узла с усиками следует один узел без усика. При росте побегов сеянцев винограда отмечается та же закономерность, но здесь чередование моноподиального роста с симподиальным осуществляется выше по длине побега, вследствие чего первый усик появляется обычно после образования 6—10-го междоузлия.
Филиппенко (1957) считает, что заложение первого усика на побеге сеянцев винограда является показателем их готовности к вступлению в плодоношение.
Несмотря на проведенные исследования, вопрос об отношении сеянцев к длине дня не вполне выяснен, что вызвало необходимость в дополнительном изучении (Стоев, Занков, 1962), с целью лучшего установления влияния фотопериодического воздействия на характер роста и развития сеянцев винограда.
В табл. 8 представлены данные о влиянии фотопериодического воздействия на рост и вызревание побегов и на заложение соцветий. В качестве наиболее существенного вывода из этих исследований можно отметить, что фотопериодическое воздействие в течение 6 и 8 h подавляет рост побегов, притом тем сильнее, чем продолжительнее это воздействие. Как правило, усики не закладываются до конца фотопериодического воздействия, но после перехода к нормальному освещению они появляются на побегах в вариантах с продолжительностью фотопериода 40 и 60 дней.
Таблица 8
Рост сеянцев сорта Памид при различной длине дня
В конце фотопериодического воздействия
В конце вегетационного периода
развитых узлов, шт.
общий прирост побегов, cm
развитых узлов, шт.
При длине дня 10 h происходит заложение усиков, но все же наблюдается подавление роста, особенно в вариантах с более продолжительным фотопериодическим воздействием (70 дней). У побегов в этих вариантах не наблюдается усиления роста и после перехода к нормальному освещению. Длина дня 12 h является нижней границей фотопериода виноградной лозы. При этой продолжительности дня во всех вариантах отмечается заложение усиков, причем подавление роста не очень сильное.
Сеянцы, подвергнутые фотопериодическому воздействию на протяжении 6—8 и 10 h, не вступают в плодоношение на второй год жизни, несмотря на то, что некоторые из них к концу фотопериода уже успели сформировать усики на побегах и в конце вегетационного периода, как правило, все сеянцы перешли к симподиальному росту. Из сеянцев, выращивавшихся при длине дня 12 h, на второй год в плодоношение вступали лишь отдельные растения, в то время как значительное число сеянцев, выращивавшихся при длине дня 18 h и при естественном освещении, вступили в плодоношение (примерно 44 и 58,8 %). Принимая во внимание то, что сеянцы, выращивавшиеся в течение первого вегетационного периода при продолжительности освещения 18 h и естественной длине дня, массово перешли к симподиальному росту, но только половина их плодоносила на второй год жизни, следует сделать вывод, что появление усика не является указанием заложения соцветий в почках, хотя растение и находится в готовности к плодоношению.
Такой вывод напрашивается тем более при сопоставлении данных о заложении усиков у растений, выращиваемых при продолжительности освещения 10—12 h, с процентом плодоносивших сеянцев в этих же вариантах.
Согласно современным взглядам, усик и соцветие имеют одинаковое генетическое происхождение. Усик по Баранову (1946) является видоизмененным соцветием. Будет ли в почках заложено соцветие или же усик, зависит, как отмечалось и выше, от условий питания в период формирования и роста зимующих почек, а также от условий следующего года во время набухания почек и начальных этапов роста молодого побега. Вопрос о существовании критических периодов (Негруль, 1952), во время которых в почке происходят существенные изменения и в результате которых она становится плодоносной или же бесплодной, рассматривался довольно подробно выше.
При анализе данных о росте побегов и формировании усиков устанавливается определенная взаимосвязь между высотой побегов, на которых появляются усики, и продолжительностью освещения. Развивающиеся в условиях более короткого дня сеянцы закладывают первые усики, как правило, выше. Например, у сеянцев, выращиваемых при длине дня 10 h, первые усики обнаруживаются на 15—16-м узле; при 12 h — на 13—15-м узле, в то время как при естественном освещении и при продолжительности освещения до 18 h усик закладывается на 11—13-м узле. Видно, что при выращивании сеянцев в условиях короткого дня, моноподиальное развитие более продолжительно, тогда как в условиях естественного освещения сеянцы быстрее переходят от моноподиального роста к симподиальному, а, следовательно, и к готовности к плодоношению.
Высота заложения первого усика, с другой стороны, находится в прямой зависимости от интенсивности роста сеянцев. Чем сильнее рост побегов в начальных фазах развития, тем ниже закладывается первый усик.
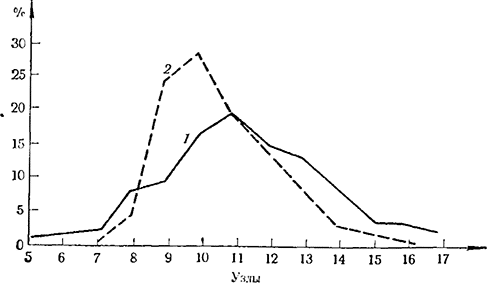
Рис. 7. Закладка первого усика у сеянцев (%)по длине побега
1—данные по 249 сеянцам (1962); 2 — данные по 963 сеянцам (1963)
Следовательно, при интенсивном росте побегов сеянцы быстрее переходят к симподиальному росту, а поскольку короткий день замедляет рост в первых фазах вегетации, становится понятным, почему растения, выращиваемые при 6—8 и 10-часовом дне, закладывают первый усик значительно выше.
Дополнительные исследования Иванова с сотр. (1968) показали, что при выращивании в условиях высокого агрофона и естественного освещения
сеянцы закладывают первый усик в секторе 10—11-го узла. На рис. 7 приведены результаты двухлетних наблюдений авторов над 1212 сеянцами, полученными от восьми гибридных комбинаций. Устанавливается, что первый усик закладывается в секторе 5—15-го узла побега, причем у преобладающей части растений (около 80%) усик обнаруживается в секторе 9—12-го узла.
Почти аналогичный характер имеют результаты Занкова (1967). Он установил, что наиболее низко находящийся узел, на котором закладывается усик, — это сектор 9—11-го узла, а наиболее высоко расположенный — в секторе 14—17-го узла. Если, однако, прищипыванием вызывается формирование пасынков первого порядка, то заложение усиков на них происходит значительно ниже. При этом усик находится тем ниже, чем выше сформирован пасынок. Еще ниже закладываются усики на пасынках второго порядка, причем они также зависят от метамерного положения пасынка.
Кроме роста надземной части, продолжительность дня оказывает влияние также на рост корневой системы. Как показали опыты Хакбарта и Шерца (цит. по Мержаниану, 1939), Потапенко и Захаровой (1937), при коротком дне рост побегов уменьшается, а усиливается относительное нарастание корневой системы. Исследования Стоева и Занкова (1962) подтверждают это явление (табл. 9).
Таблица 9
Соотношение между приростом надземной части и корневой системой (в свежем весе) сеянцев сорта Памид после 70-дневного фотопериодического воздействия
Средний вес одного побега, g
Средний вес корней, (g)
Отношение веса побегов к весу корней
Естественный день (контроль)
Вес корней сеянцев, выращиваемых при длине дня 6—12 h, в 4,0—8,8 раз больше веса побегов. У сеянцев, выращиваемых при длине дня 10 h, это соотношение наиболее велико (8,8 раза), а при естественном освещении и продолжительности дня до 18 h оно доходит до 2,3—2,6 раза.
Фотопериодическое воздействие оказывает влияние на рост сеянцев и на следующий год. Как правило, прирост сеянцев, развивающихся при коротком дне, в следующем году слабее по сравнению с сеянцами, развивающимися при естественном освещении. В соответствии с этим число развивающихся узлов и толщина побегов у основания также меньшие.
Физиология прорастания семени
Покой семян. Семена винограда, как и ряда других растений, имеют период покоя. Глубина и продолжительность покоя семян V. vinifera и остальных видов винограда изучены мало. В семенах растений содержатся вещества, которые подавляют прорастаемость (триптофан). В том, что семена винограда проходят фазу покоя, легко убедиться при одновременном посеве сухих и стратифицированных семян. К аналогичным выводам можно прийти и при сопоставлении прорастаемости семян, не прошедших предварительной предпосевной обработки, но при посеве в различные сроки. Пребывание семян в почве на протяжении некоторого периода при оптимальных сроках сева и наличии соответствующих условий (влажность, температура и аэрация), по всей вероятности, оказывает влияние, аналогичное предпосевной подготовке, в результате чего их всхожесть повышается.
Для повышения прорастаемости семян, получения быстрых дружных всходов и выравненных сеянцев большинство ученых рекомендуют предварительную стратификацию семян винограда.
Неодинаковы мнения различных специалистов также относительно продолжительности стратификации. Одни считают, что стратификация должна продолжаться 180–210 суток при температуре от 1 до 3°С. Такой срок предпосевной обработки необходим для разрушения оболочки семян, которая считается главным препятствием для прорастания зародыша. Продолжительная стратификация, благоприятно влияет на рост и вызревание побегов, а также на успешную зимовку сеянцев.
Стратификация семян должна начинаться сразу после их извлечения из ягод и проводиться при температуре от 0 до 7°С, 90-120суток.
Установлена определенная зависимость между общим процентом проросших семян, динамикой прорастания и температурой, при которой проводилась стратификация, с одной стороны, и продолжительностью стратификации семян при одной и той же температуре, с другой. Как правило, семена, стратифицированные при температуре ниже 0°С (от –5° до –1°С), имеют всхожесть меньше, чем стратифицированные при температуре выше 0°С (от 5° до 10°С). Кроме того, прорастание первых начинается и заканчивается позднее. В конечном итоге оказывается, что для стратификации семян при температуре ниже 0°С необходимо более продолжительное время. Например, при –5°С наиболее высокой всхожести семена достигают через 120 суток, а при 5°С — через 90 суток.
Следует отметить, что продолжительность стратификации семян неодинакова. Как при –1°С, так и при положительных температурах (5 и 10°С) через 30 дней способность прорастать приобрело более 65% семян. Семена некоторых сортов имеют сравнительно короткий период покоя (менее 30 дней), поэтому в более холодные и влажные годы они не обнаруживают состояния покоя. Если замочить такие семена в течение 5–7 суток, они прорастут на 70–80%. Семена других сортов имеют период покоя продолжительнее и требуют более длительного периода стратификации (70–120 суток).
Для повышения прорастаемости семян винограда и стимуляции роста сеянцев некоторые авторы предлагают и другие методы, применяемые отдельно или совместно со стратификацией.
Высокий процент проросших семян винограда и сильное развитие сеянцев в течение первого года, получается при замачивании семян до стратификации в 1- и 3%-ном растворе Na2,C03, и 0,25- и 3%-ном растворе (NH4)2S04 и меди (15–20 мг на 150 м3 воды).
Также сильно увеличивает прорастаемость семян винограда обработка NaHC03, NaCl, молочной и уксусной кислотой. Кроме того, наблюдается более сильное развитие сеянцев в первый вегетационный период. Аналогичные результаты получаются при обработке семян винограда до стратификации никотиновой, масляной и индолилмасляной кислотой в различных концентрациях.
По данным некоторых авторов, положительное влияние на прорастание семян оказывает комбинированная обработка, при которой семена после стратификации в течение 30–60 суток проращивают, а затем снова стратифицируют при различной температуре.
Прорастаемость семян. Условия, определяющие непрорастаемость семян винограда, не вполне выяснены. Некоторые авторы объясняют непрорастаемость семян непроницаемостью семенных оболочек (кожуры семян).
Процентная разница всхожести скарифицированных и обычных семян у некоторых сортов небольшая, а у некоторых значительно выше. Во всех случаях, однако, скарификация не приводила к повышению содержания воды в семенах, как это предполагалось.
Значительное влияние на всхожесть семян оказывает температура. Сорта винограда делятся по их требованиям к температуре на две группы: тип винифера, для которых оптимальная температура около 30°С, а минимальная и максимальная–20–35°С; тип солонис, для которых оптимальная температура около 27°С, с более широким интервалом между минимумом и максимумом.
По неопубликованным данным, для прорастания семян ряда сортов оптимальная температура 28°С. Существуют, однако, и значительные отклонения, выявляющие неодинаковые температурные требования сортов V. vinifera. Для некоторых сортов оптимальная температура значительно более низкая (около 20°С). Например, около 38% семян одного сорта прорастали при температуре 15°С, а у других сортов, всхожесть семян при температуре 15°С незначительная (от 1,5 до 4,5%).
При температуре 35° всхожесть семян сортов (с высокой прорастаемостью при 15°С), уменьшается незначительно, тогда как у других сортов заметно. При температуре выше 40°С семена всех сортов теряют способность прорастать. При 8–10°С начинают прорастать лишь единичные семена, что дает основание предполагать, что самая низкая для прорастания семян винограда температура около 10°С.
Мнения различных авторов о сроках извлечения семян из ягод винограда и способах их хранения до начала предпосевной подготовки не совпадают. Большинство специалистов придерживаются мнения, согласно которому, полностью созревшие грозди нужно хранить в сухих, вентилируемых и холодных помещениях до конца осени.
При хранении семян до посева в ягодах всхожесть семян уменьшается значительно, причем тем сильнее, чем продолжительнее хранение. В процессе хранения ягоды постепенно теряют воду, в них резко уменьшается газообмен, что в конечном счете приводит к снижению всхожести семян. Поэтому семена следует извлекать из ягод не позже двух месяцев после сбора винограда и сразу же стратифицировать.
Также отрицательно влияние длительного хранения семян в воздушносухом состоянии на их жизнеспособность. Основной причиной снижения всхожести семян – уменьшение их влажности. После хранения на протяжении 12 месяцев семена потеряли 30,2% воды.
В заключение можно сказать, что для селекционных целей семена нужно собирать после наступления полной физиологической зрелости ягод. Дальнейшее оставление гроздей на кустах не повышает всхожести семян. Извлекать семена из ягод можно сразу после сбора винограда, или несколько позднее, спустя 2–3 месяца, если виноград хранится в условиях, не вызывающих его повреждений или загнивания. После подсыхания до воздушносухого состояния (10– 15 дней на фильтровальной бумаге) семена нужно хранить в комнатных условиях до начала предпосевной подготовки.
Источники:
http://vinograd.info/info/fiziologiya-vinograda/fiziologiya-razmnozheniya-vinograda-semenami.html
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/fiziologiya-rosta-i-razvitiya-semyan-i-seyancev-vinograda-9.html
http://m.studwood.ru/1973881/agropromyshlennost/fiziologiya_prorastaniya_semeni





