
Красящие вещества (антоцианы) – Основные закономерности роста и созревания ягод винограда
Красящие вещества (антоцианы) – Основные закономерности роста и созревания ягод винограда
Красящие вещества (антоцианы) – Основные закономерности роста и созревания ягод винограда




Характеристика биохимических процессов роста и созревания винограда
Со времени образования завязи основная энергия виноградного куста направляется на развитие ягод, что немедленно сказывается на уменьшении прироста побегов лозы.
По характеру изменений, происходящих при развитии виноградной ягоды, можно различать три периода: 1) роста; 2) созревания и 3) перезревания.
Период роста начинается с образования завязи и продолжается до начала размягчения ягоды и появления у красных сортов окраски; продолжительность этого периода 40-45 дней. В этот отрезок времени происходит энергичный процесс дыхания, пластические вещества расходуются на построение клеток и тканей ягоды, которая постепенно увеличивается в размерах. Из семяпочек образуются семена, околоплодник разрастается. Ягода содержит хлорофилл, окрашена в зеленый цвет и так же, как и другие зеленые части растения, усваивает углекислоту воздуха. Однако образующийся сахар расходуется, накопления его не происходит, вследствие чего содержание его в ягодах в этот период не больше, чем в других зеленых органах виноградной лозы и редко превышает 1%. Сахар в виноградной ягоде представляет собой преимущественно глюкозу (около 75%).
Во время роста ягоды обогащаются органическими кислотами и их кислыми солями.
Органические кислоты в виноградной ягоде образуются, как и в листьях, в результате сгорания сахара. Вначале появляется большое количество свободной яблочной и винной кислоты. По мере развития ягод содержание кислот возрастает и кислотность сока достигает нередко 35 г/л (на винную кислоту) и выше. Яблочная кислота все время преобладает над винной и к концу периода роста составляет главную часть кислот в соке ягод. Винного камня в этот период сравнительно мало. Кроме яблочной и винной, в виноградном соке в незначительных количествах встречаются кислоты лимонная, янтарная, гликолевая, щавелевая, глюкуроновая и другие.
Общее количество дубильных веществ в ягоде постепенно возрастает, по в соке мякоти к концу периода роста количество их заметно падает.
Несмотря па быстрый рост клеток, зеленые ягоды до конца периода остаются твердыми. Увеличение объема мякоти ягоды сопровождается также ростом семян, вес которых в течение всего периода увеличивается, причем они остаются зелеными и мягкими.
Таким образом, характерной особенностью периода роста является накопление в ягоде органических кислот, образующихся в результате окисления углеводов.
Период созревания. К концу первого периода ягода перестает расти и достигает нормальной величины. Если в дальнейшем ягода и увеличивается, то очень незначительно. Переход плодовой мякоти из твердого в мягкое состояние (превращение протопектина в пектин) создает впечатление, будто ягода наливается соком. Поэтому период созревания называют также периодом налива ягод. Помимо размягчения у красных сортов появляется красная окраска, у белых – светло-желтая или желто-зеленая.
С прекращением роста ягоды перестают расходовать углеводы, вследствие чего в них быстро накапливается сахар за счет поступления его из листьев. Это накопление сахара так велико, что дневной прирост его достигает 0,5-1%, особенно в конце периода созревания. При этом наряду с глюкозой накапливается фруктоза, количество которой постепенно относительно возрастает и к концу периода достигает той же величины, что и количество глюкозы. Полная зрелость наступает тогда, когда абсолютное накопление сахара останавливается.
Другим, не менее важным, признаком, характеризующим период созревания ягод винограда, является падение кислотности сока. Кислоты винная и яблочная испытывают в этот период различные изменения. Винная кислота нейтрализуется основаниями, преимущественно калием, образуя кислый виннокислый калий. Этот процесс совершается в течение всего периода созревания винограда и к концу его винная кислота почти целиком переходит в соли, так что в соке вполне зрелых ягод в свободном состоянии она не содержится.
Яблочная кислота, как менее активная, чем винная, значительно слабее реагирует с основаниями; количество ее уменьшается вследствие окисления до углекислоты и воды в процессе дыхания. Частично яблочная кислота переходит в другие органические соединения, а в конце периода созревания, когда свободной винной кислоты обычно уже нет, дает соли. К моменту полной зрелости ягод яблочная кислота, как показывают исследования, в небольшом количестве содержится в них в свободном состоянии.
Таким образом, количество кислот при созревании уменьшается вследствие нейтрализации их минеральными веществами, поступающими в ягоду, и окисления при дыхании до окончательных продуктов – СО2 и Н2О. Наряду с накоплением сахара и уменьшением кислот в период созревания происходит ряд других, менее изученных явлений. К числу их относятся изменения пектиновых веществ, сопровождающиеся переходом нерастворимого протопектина в пектин, в результате чего ткани мякоти размягчаются и ослизневают. Вместо хлорофилла в ягоде появляются красящие и ароматические вещества. Общее содержание азотистых веществ изменяется очень мало, но в то же время количество нерастворимых азотистых веществ возрастает. В золе все время увеличивается содержание калия, кальция и магния. Количество фосфорной кислоты также повышается. Дубильные вещества из мякоти исчезают, но общее содержание их в ягоде существенно не изменяется.
Характерный для сорта аромат проявляется только при полной зрелости ягод.
На созревание ягод оказывает большое влияние внешняя температура; оптимальная температура 30°. При более высокой температуре ягоды перегреваются, подвергаются «запалу» и задерживаются в росте. Свет зреюшим ягодам не нужен.
Важное значение во время созревания винограда имеет влага. Только при достаточном ее притоке из почвы происходит нормальная работа хлорофилла в листьях, регулярное поступление минеральных веществ и правильное распределение питательных веществ в растении. В сухую погоду из-за недостатка влаги созревание ягод сильно задерживается, что выражается в крайне медленном накоплении сахара. В засушливые годы ко времени сбора винограда процесс созревания в нем приостанавливается и в ягодах не накапливается того количества сахара, которое они обычно имеют при достижении полной зрелости.
Гребни в период роста и созревания содержат большое количество крахмала. К моменту полной зрелости винограда крахмал из гребней исчезает, они деревенеют, теряют воду и вес их значительно уменьшается. Семена также деревенеют, приобретают коричневую окраску; вес их от начала созревания до полной зрелости уменьшается.
Период перезревания. При наступлении полной зрелости гребни деревенеют, вследствие чего связь между виноградным растением и ягодами прерывается, сок ягоды начинает концентрироваться в результате испарения содержащейся в ней воды. Содержание сахара и количество сухого вещества в ягодах относительно повышается. Абсолютное же количество сахара, а также кислот уменьшается вследствие сгорания при дыхании ягод. Разрушению подвергается преимущественно фруктоза; исследования показывают, что ее относительное количество все время понижается. Потеря сахара вследствие сжигания в третий период невелика.
Кислотность в период перезревания, несмотря на концентрацию сока ягоды, в результате сжигания яблочной кислоты и нейтрализации кислот основаниями, все время падает. Кислотность сусла значительно уменьшается, что ясно ощущается на вкус.
Изменения других веществ в винограде при его перезревании мало изучены.
Основные закономерности роста и созревания ягод винограда – Красящие вещества винограда (антоцианы)
Содержание материала
В ягоде винограда, в особенности в кожице, имеется ряд веществ,-определяющих окраску ягод, а после брожения — и окраску вина. Цвет ягод и·’цвет вина не только специфический сортовой признак — он имеет существенной значение для качества винограда и вина. Поэтому к вопросу о природе красящих веществ и факторов их изменчивости всегда проявлялся большой интерес.
Красящие вещества, встречающиеся в ягоде, обычно разделяют на две группы: зеленые и желтые красящие вещества и красные красящие вещества.
К первой группе относят хлорофилл, ксантофилл и каротин (антоксантины). Они находятся в ягодах до начала созревания, после чего обнаруживаются только следы этих веществ. Пигменты, обусловливающие цвет белых сортов от светло-зеленого до золотисто-янтарного (флавоны), являются продуктами изменений хлорофилла, ксантофилла, каротина, их производных и других веществ, сопутствующих хлорофиллу (Баранов, 1946). Основную часть флавонов составляют кверцитин и кверцитрин.
Хлорофилл и желтые пигменты содержатся не только в кожице и мякоти винограда белых сортов, но также в кожице и мякоти окрашенных сортов (Фролов-Багреев, 1959; Gаrd, 1903).
К группе красных красящих веществ относят антоцианы, которые являются собственно красящими веществами в ягодах окрашенных сортов винограда. Они находятся в кожице и мякоти.
На рис. 15 приводится схематическое изображение расположения красящих веществ в кожице 7 групп наиболее типичных по своей окраске сортов (Негруль, Лю-Юй-Янь, 1963).
Первые сведения о красных красящих веществах в винограде и вине (антоцианах) появились еще в начале прошлого столетия (см. сводки Дурмишидзе, 1955; Негруль, Лю-Юй -Янь, 1963).
В конце XIX — начале XX столетия был проведен ряд исследований, среди которых следует отметить работы Gautiе (1892), Неisе (1889), Wе1gеrt (1895), Petri (1903). Самые обстоятельные исследования содержания и природы красящих веществ в винограде и вине принадлежат Willstater с сотр. (1915, 1917).
Многочисленные исследования красящих веществ в винограде и вине проведены Ribereau-Gayon (1953, 1958), J. и Р. Ribereau-Gayon (1954), Fonas sin (1956), Дурмишидзе (1955), Дурмишидзе и Нуцубидзе (1958), Дурмишидзеи Сопромадзе (1963), Негрулем и Лю-Юй-Янь (1963), Кондо и Звездиной (1953), Бегуновой (1953), Стуруа с сотр. (1973), Abbасh с сотр. (1959), Rankin с сотр. (1958), Маркосовым (1973) и многими другими. Благодаря этим исследованиям теперь хорошо известно, что антоциан винограда (энин) представляет собой моноглюкозид энидина, а энидин является диметиловым эфиром дельфинина.
При изучении антоцианов винограда разных видов р. Vitis установлено значительное разнообразие красящих веществ. Поэтому сейчас уместнее говорить об антоциановом комплексе, который у разных сортов и видов не имеет однородного состава. Главный компонент пигмента у сортов — V. vinifera — энин, а у американских — ампелопсин.
Исследованиями Bate-Smit и Westal (1950), Ribereau-Gayon (1953, 1958), J. и Р. Ribereau-Gayon (1954), Дурмишидзе с сотр. (1955, 1958) и др. установлено, что красящие вещества винограда европейских сортов состоит в основном из моноглюкозидов мальвидина, петунидина, дельфинидина, цианидина и пеонидина. В составе красящих веществ винограда американских видов и европейско-американских гибридов имеются в основном диглюкозиды указанных аглюконов, причем комплекс антоцианов у разных сортов значительно изменяется. Как отмечают Дурмишидзе и Нуцубидзе (1958), комплекс пигментов может содержать от одного до десяти и более антоцианов.

Рис. 15. Поперечный (а, б, в, г) и продольный (д, е, ж, з) срезы кожицы ягод винограда следующих сортов
а — Саперави (тип 1); б — Кара узюм ашхабадский (тип 2); в — Додреляби (тип 3); г — Каш узюм (тип 5)
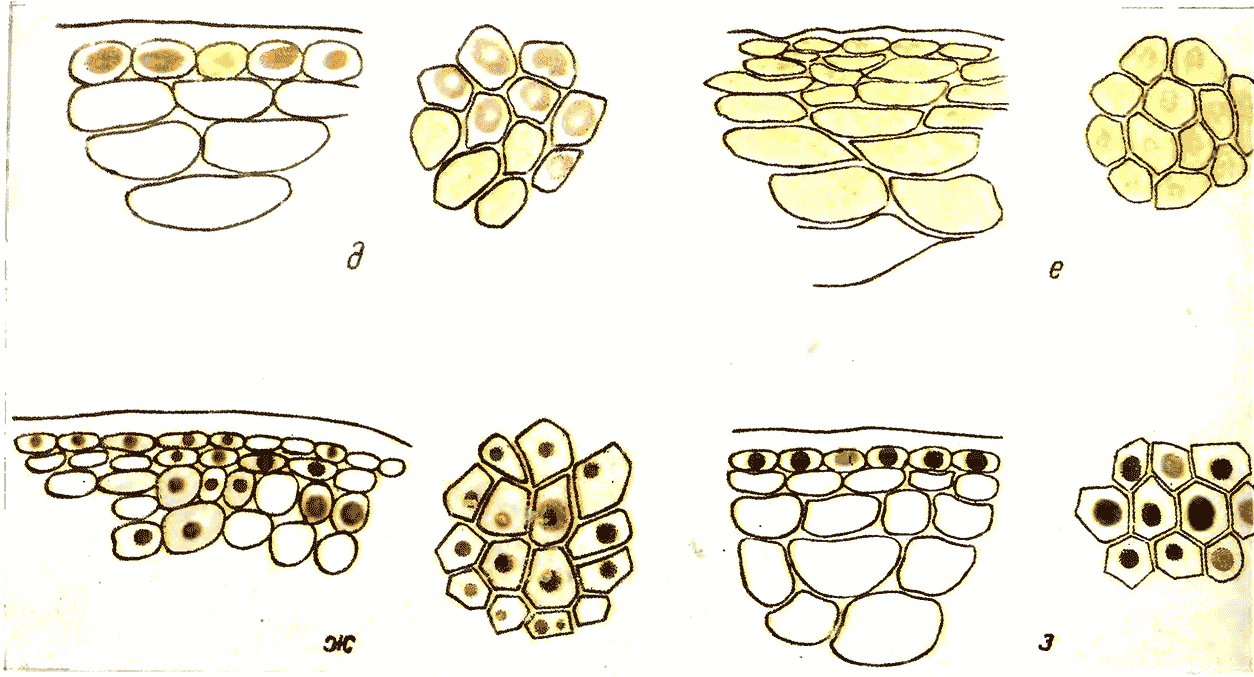
Рис. 15. Поперечный (а, б, в, г) и продольный (д, е, ж, з) срезы кожицы ягод винограда следующих сортов д —Сары Ангушты (тип 6); е — Новат нияз Халили (тип 7); ж — Зогак (тип 10); з — Марокен серый (тип 12)
Например, в винограде сорта Цители- Ркацители авторы нашли один антоциан — моноглюкозид мальвидина, а в кожице винограда сорта Алеатико — одиннадцать. Аналогичные выводы сделаны в работе Гетова (1966, 1966а, 1969).
Дальнейшие исследования красящих веществ в ягоде винограда были направлены, главным образом, на изучение специфического характера антоцианов у различных видов р. Vitis, влияния экологических и метеорологических факторов на содержание антоцианов и наследование пигмента типа Мальвина. По некоторым из этих вопросов на протяжении двадцати лет ведется оживленная дискуссия.
Одним из основных направлений изучения красящих веществ, по которому различные исследователи еще не достигли единой точки зрения, является вопрос о содержании диглюкозидов в ягодах сортов V. vinifera. Ribеrеаu- Gayon и Sudraud (1957) утверждают, что моноглюкозиды специфичны только для сортов V. vinifera, в то время как диглюкозиды находятся в главнейших видах р. Vitis. Основные выводы из этих исследований приведены в табл. 13.
Таблица 13
Состав красных красящих веществ в ягодах, %
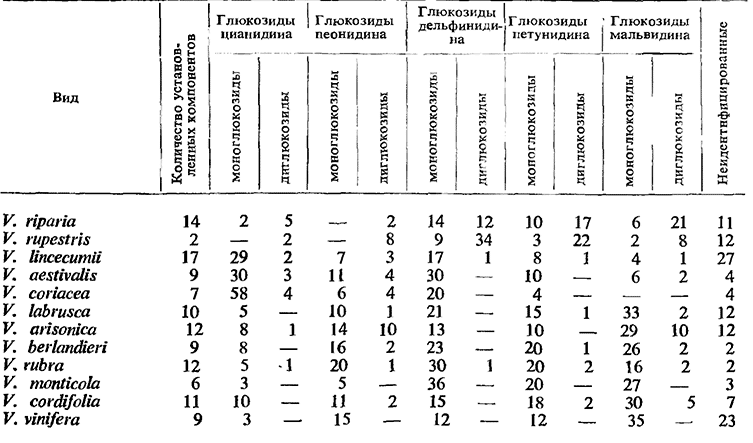
Из данных таблицы видно, что в ягоде винограда имеется большое число пигментов (до 17), притом у различных видов их состав не идентичен. Наиболее характерно то, что у сортов V. vinifera не обнаружено диглюкозидов. Это дало основание Ribereau-Gayon (1959) сделать вывод, что содержание диглюкозидов является специфическим ботаническим показателем. Этим автором было доказано также, что в винах, полученных из винограда сортов, относящихся к виду V. vinifera, нет диглюкозидов, в то время как в винах из винограда американских сортов или же гибридов диглюкозиды всегда имеются (Ribereau-Gayon, Peynaud, 1960). На этой основе был разработан метод контроля происхождения вин и установления наличия в них винограда гибридов (Ribereau-Gayon, 1954, 1956).
Проведенные в ряде других лабораторий исследования, однако, не подтвердили выводы Ribereau-Gayon о специфическом характере антоцианов в ягодах сортов V. vinifera, что вызвало на V Международном конгрессе биохимиков оживленную дискуссию по этому вопросу.
Дурмишидзе и Нуцубидзе (1958) установили, что у ряда сортов V. vinifera грузинского, итальянского и португальского происхождения имеются диглюкозиды дельфинидина, петунидина и мальвидина.
Возможность наличия диглюкозидов в ягодах вслед за Дурмишидзе признают Deibner и Bourzeix (1960, 1968). Диглюкозиды антоцианов в ягодах V. vinifera обнаружили также Нудубидзе иГулбани (1959).
Малышева и Кострикин (1973) исследовали 220 сортов на содержание мальвидина. Из европейских сортов они установили наличие диглюкозида мальвидина только у шести: Ширван шахи, Хиндогни, Мускат черный (Кальяба), Араксени черный (Езандари), Марсельский ранний и Ян изюм черный. В то же время авторы не обнаружили диглюкозида мальвидина у таких сортов, как ВИР-1, Матраса, Мускат гамбургский, Португизер и др. у которых, по утверждению других авторов, он имеется. У всех европейско- амурских гибридов мальвин доминирует, однако при возвратном скрещивании возможно получение сеянцев, сочетающих высокую морозостойкость отсутствием мальвина. Очень важен вывод авторов, что диглюкозид не появлялся и не исчезал под влиянием экологических условий.
Коадэ (1971) исследовал пигменты в кожице винограда 12 видов рода Vitis, 7 разных сортов вида V. vinifera и 5 сортов прямых производителей. Полученные результаты показали, что у исследованных сортов V. vinifera не имеется мальвина, а у всех остальных видов рода Vitis и сортов прямых производителей он обнаруживается (табл. 14).
Кострикин и Малышева (1972) исследовали 89 сортов V. vinifera. Из них мальвин они установили только у винограда сорта Хиндоги и Мускат черный (Кальяба). Мальвин нашли также у V. amurensis, а из гибридов с участием амурского винограда — у сортов Степной, Саперави северный, Фиолетовый ранний и Фестивальный. У V. vinifera sylvestris мальвин не установлено. У амурского винограда Грицун (1970) также установи, наличие диглюкозида мальвидина.
Негруль и Лю-Юй-Янь (1963) изучали пигментный состав 5. сортов винограда в условиях Средней Азии (Ташкент) и 53 сортов в условиях южного берега Крыма (Ялта), относящихся к разным типам, по принятой ими классификации сортов по окраске ягод. Из данных, приведенных в табл. 15, видно, что разная окраска ягод винограда обусловлена различным содержанием пигментов и особенностями их сочетания в кожице и соке.
Видно также, что у сортов с темно-синей, темно-красной, темно-коричнево-красной, фиолетовой и красной окраской ягод обнаружены диглюкозиды типа мальвидина. На этом основании авторы заключают, что резкого различия в содержании антоцианов и ягодах европейско-азиатского и американского винограда нет.
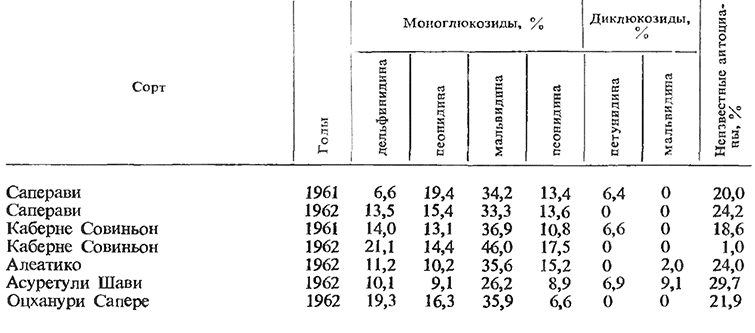
В более поздней работе Дурмишидзе и Сопромадзе (1963, снова сообщают, что они обнаружили в ягодах некоторых сортов V. vinifer наличие диглюкозидов петунидина и мальвидина (табл. 16).
Гетов, Петков (1966) при изучении антоцианового комплекса 19 сортов ампелографической коллекции Института виноградарства обнаружил наличие диглюкозида мальвидина у следующих сортов: Дамас нуар, Кравцици, Марсильский черный ранний, Матраса, Мускатный Магарач, Пармак черен, Пармак червен, Плавина мала, Прюн дю Казул, Станушина, Хиндогни, Хый зи син, Черно едро, Чхавери.
Таблица 14
Качественная характеристика антоцианов кожицы ягод некоторых видов и сортов винограда (1969 г.)

Условные обозначения: дл — дельфинидол; пт — петунидол; мл — мальвидол; пн — пеонидол; X1, X2, Х3, Х4 — неидентифицированные (ацилированные) пигменты; + наличие пигментов; — их отсутствие.
Окраска всех сортов черная.
Авторы заключают, что либо не имеется резкой границы в антоциановом комплексе между сортами V. vinifera и сортами — прямыми производителями, или же сорта, у которых обнаружен диглюкозид мальвидина, являются гибридами с участием американских сортов.
Далее Петков (1977) изучал динамику и содержание антоцианового комплекса у сортов Букет, Руен и Рубин в двух экологических районах на протяжении трех лет (табл. 17). Из антоцианов установлены только моноглюкозиды дельфинидина, петунидина, мальвидина и пеонидина. Кроме того, на хроматограммах установлено наличие 4 неидентифицированных антоциановых веществ.
Многочисленные исследования проводили Cappelleri с сотр. (1964), Cappelleri (1964, 1965, 1965а), Liuni (1965), Cappelleri (1966, 1966а), Cappelleri (1967) в целях установления присутствия диглюкозидов в антоциановых пигментах разных сортов вин и изучения их состава или же для разработки методики.
Таблица 15
Содержание различных пигментов в ягодах винограда, относящихся по окраске к разным шкалам
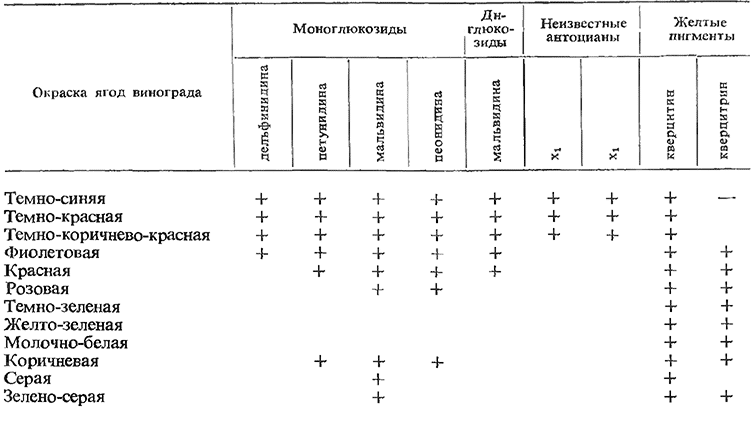
Таблица 16
Состав антоцианов в кожице разных сортов винограда (местность Дигоми)
Они установили диглюкозид мальвидина в ягодах сортов Мерло, Каберне фран, Рабозо ди Пиаве и Мускат гамбургский. Диглюкозид мальвидина они установили также в вине из сортов Неграра ди Веронезе и Негронца. В ягодах же ряда гибридов прямых производителей и некоторых подвоев мальвидин не установлен. Ими не установлено также специфического комплекса антоцианов у разных видов рода Vitis.
Дальнейшие исследования Сарре11еri (1970) показали, что диглюкозидов, в том числе диглюкозидов мальвидина, не имеется в вине ряда прямых производителей с розовой окраской ягод (Galibert 21-2, Galibert 22-4, Seibel 7053, Seibel 10878, Seibel 13666, Seibel 14596).
Содержание антоциановых веществ в кожице созревших ягод винограда (% от абс. сух. вещества)
Основные закономерности роста и созревания ягод винограда
Содержание материала
К. Д. Стоев
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ РОСТА И СОЗРЕВАНИЯ ЯГОД ВИНОГРАДА
При изучении физиологии органов виноградной лозы большое внимание уделено гроздям и ягодам винограда. Это вполне понятно, так как ягода винограда — единственный орган, имеющий хозяйственное значение. Благодаря высокому содержанию сахаров и других полезных органических и минеральных веществ, она является ценнейшим плодом для потребления в свежем виде или же для промышленной переработки и получения виноградных вин и других напитков. Поэтому в течение многих десятилетий изучались рост гроздей и ягод винограда и накопление в них ценных питательных веществ. На протяжении столетий на основе многочисленных эмпирических наблюдений вырабатывалось понятие о качестве винограда и обусловливающих его факторах. Пожалуй, нет другой сельскохозяйственной культуры, о качестве продукции которой накопилось бы столько сведений и к которой предъявлялись бы такие высокие требования, как к винограду и продуктам его переработки.
КРАТКИЕ СВЕДЕНИЯ ОБ АНАТОМИЧЕСКОМ СТРОЕНИИ ГРОЗДЕЙ И ЯГОД
По своему анатомическому строению кисть винограда напоминает молодой побег, а впоследствии и вызревший.
Ягода винограда состоит из эпикарпия (кожицы), мезокарпия (мякоти) и эндокарпия. Кожица составлена из эпидермиса и 10—15 слоев клеток, расположенных под ним. Строение кожицы, в т. ч. толщина, плотность, количество составляющих ее клеточных слоев, а также толщина и состав кутикулы специфичны для отдельных сортов и в значительной степени определяют устойчивость ягод против Botritis Cinerea, транспортабельность и лежкость винограда.
Кожицу делят на две подзоны. Наружную, состоящую из 2 до 8-9 узких, тангентально удлиненных и утолщенных стенок, обозначают знаком х1 (первая подзона гиподермиса). Внутренняя подзона охватывает также несколько слоев (Кодряну, 1976), расположенных в тангентальном направлении. Клетки внутренней подзоны значительно крупнее.
Количество клеточных слоев, их толщина и плотность в первой подзоне неодинаковы у разных сортов и варьируют, по данным Яноша (цит. по Карадимчевой, 1980), от 40—50 до 170—180 μ, а по данным Кодряна —
в пределах между 40 и 100 μ. Исследования Карадимчевой (1980) показали, что толщина слоя находится в пределах 50—120 μ, причем у сортов Гымза, Алиготе и Чаушона составляет 50—70 μ, а у сортов Каберне Совиньон, Мерло, Болгар и Уньи белый — 100—120 μ. Считается, что именно количество и толщина клеточных слоев этой подзоны обуславливает устойчивость ягод к загниванию.
Исследования показали также, что толщина кутикулы мало изменяется по годам (табл. 1):
Таблица 1
Толщина гиподермального слоя х1 (μ)
Эпидермис состоит из одного слоя клеток толщиной 30—40 μ. Сверху эпидермальные клетки утолщены и кутинизированы. Толщина кутикулы специфична для отдельных видов и, по данным Bonnet (1903), варьирует в следующих пределах: V. vinifera — 1,5—3,8 μ; V. rupestris — 4,1—4,6 μ; V. berlandieri—8,1—8,5 μ; V. coriacea — до 10 μ.
Мякоть (мезокарпий) составляет наибольшую часть ягоды. Она состоит из 11—16 слоев крупных клеток паренхиматического строения. Каждая клетка имеет тоненькую мембрану и заполнена клеточным соком — соком винограда. Мембраны клеток мякоти ягод не прилегают плотно друг к другу, между ними имеются небольшие межклеточные, сообщающиеся между собой пространства, посредством которых осуществляется газообмен с внешней средой. В мякоти выделяются две зоны. Первая находится непосредственно под кожицей и состоит из нескольких слоев (расположенных радиально), которые достигают значительной величины — до 0,3—0,4 mm в диаметре. Мембраны этих клеток очень тонкие (от 0,5 до 1 μ) и часто во время созревания ягод размягчаются. Вторая зона состоит из клеток, большая часть которых также расположена радиально. Их мембраны несколько толще и не размягчаются. Самый внутренний, прилегающий к семенам слой мякоти составляет эндокарпий. Он также состоит из радиально расположенных клеток.
Общее количество составляющих ягоду винограда клеточных слоев от эпидермиса до эндокарпия включительно — около 25—30, столько же, сколько у завязи. Таким образом, увеличение объема ягоды вызвано нарастанием элементов, а не увеличением их числа.
Исследования Мержаниана (1940) показали, что на первом этапе роста ягоды деление клеток происходит в тангентальном направлении, главным образом с момента оплодотворения до достижения ягодами величины
2—3 mm в диаметре. В дальнейшем при росте ягоды количество клеток не увеличивается. По данным А. С. Мержаниана, количество клеточных слоев мякоти остается весьма близким к количеству клеточных слоев кожицы в течение всего роста ягоды.
У столовых сортов винограда клетки 6-7 периферических слоев, которые вместе с эпидермисом составляют кожицу ягоды, во время созревания сильно увеличиваются к внутренности мякоти. Их мембраны имеют такую же плотность, как и в первой зоне мезокарпия. Таким образом у ягод столовых сортов переход от кожицы к зоне мезокарпия осуществляется без каких-либо существенных анатомических различий. Мембраны этих клеток выделяют небольшое количество сока, что обусловливает характерную мясистую консистенцию мякоти. Отсутствие анатомического разграничения между кожицей и зоной мезокарпия приводит к тому, что они составляют единое целое, которое легко раздавливается, без отделения кожицы.
У технических сортов клеточных слоев кожицы меньше, клетки мельче, с более толстыми мембранами, чем у клеток первой зоны мезокарпия. Поэтому кожица ягоды винных сортов заметно отграничена от мякоти и чаще всего отделяется при раздавливании ягоды. Мезокарпий же составлен из значительно более крупных клеток, с очень нежными мембранами, которые при созревании винограда растворяются, в результате чего в ягоде под кожицей имеется сплошная жидкая зона.
Описанные подробности анатомического строения ягод столовых и винных сортов характерны для наиболее типичных представителей обеих групп сортов. Между ними существуют все степени перехода из одной группы в другую.
Источники:
http://wine.historic.ru/books/item/f00/s00/z0000015/st023.shtml
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/osnovnye-zakonomernosti-rosta-i-sozrevaniya-yagod-vinograda-6.html
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/osnovnye-zakonomernosti-rosta-i-sozrevaniya-yagod-vinograda.html




