
Изменения содержания гемицеллюлозы в процессе роста и вызревания побегов – Стеблевые части
Изменения содержания гемицеллюлозы в процессе роста и вызревания побегов – Стеблевые части
Стеблевые части – Дополнительные данные о вызревании побегов винограда
Содержание материала
Ж. Буар
ДОПОЛНИТЕЛЬНЫЕ ДАННЫЕ О ВЫЗРЕВАНИИ ПОБЕГОВ
Вызревание побегов соответствует совокупности морфологических, анатомических и биохимических изменений, в ходе которых побеги переходят из травянистого состояния в одревесневшее. Во время листопада вызревание завершается. В этот период побеги уже вызрели, но не в одинаковой степени.
Случай боковых побегов, или пасынков несколько особый в том смысле, что их вызревание, по-видимому, связано с потенциальными возможностями почек, из которых они развиваются, иными словами, оно зависит от ритмических изменений роста, совершающихся в лозе. Как известно, очередные пазушные почки типов Pb0, Pb1 и Рb2, расположенные соответственно на уровне узла, у которого нет усика, и следующих двух узлов, у которых усики имеются, обладают неодинаковыми возможностями развития. У сорта Уньи белый (Воuаrd, 1966) пасынок, образовавшийся из Рb0, развивается сильнее двух последующих пасынков, вызревает лучше, чем пасынки, образовавшиеся из Рb2 и особенно из Рb1 и является единственным, остающимся зимой на главном побеге, тогда как Pb1 исчезает полностью, а Рb2 в большинстве случаев представлен одним или двумя междоузлиями.
Новые данные в связи с вызреванием относятся только к его биохимической стороне: они касаются не только углеводов, но и других компонентов побега — органических кислот, аминокислот, жирных кислот, феноловых соединений. Эти данные получены в результате систематических исследований побегов множества сортов винограда в течение всего годичного цикла на протяжении многих лет, с распускания почек до следующего распускания. Их можно обобщить следующим образом.
ИЗМЕНЕНИЯ НЕРАСТВОРИМЫХ И ОБЩИХ УГЛЕВОДОВ В ТЕЧЕНИЕ ГОДИЧНОГО ЦИКЛА
Углеводы виноградной лозы, в особенности крахмал, являлись предметом многочисленных исследований (Kovessi, 1901; Vidal, 1911; Winkler, Williams, 1945; Стоев, 1947—1948; Стоев, 1952; Еifеrt с сотр., 1961). Общие углеводы (GT), о которых здесь идет речь, представляют собой совокупность общих растворимых углеводов (GST), редуцирующих (GR) и нередуцирующих (GNR) и нерастворимых углеводов (GI). Последние отвечают углеводам, полученным в результате кислотного гидролиза (при помощи НС1 2N) измельченного вещества, оставшегося после извлечения GST и размолотого до степени соответствия размеров его частиц нормам установленным Стоевым (1952), т. е. не крупнее 0,25 mm.

Рис. 1. Изменение содержания углеводов (%) в основании побегов Рупестрис дю Ло (В о и а г d, 1966)
GT — общие углеводы; GI — нерастворимые; GST— общие растворимые; GNR — нередуцирующие; GR — редуцирующие углеводы
Графики, относящиеся к междоузлиям у основания побега (рис. 1), ясно показывают, что GI и GT (кстати, то же самое касается и GST, GR и GNR претерпевают непрерывные изменения в течение годичного цикла как в период покоя (после листопада), так и в период активной жизни. Иными словами, на полученных кривых, соответствующих трем фазам развития побегов, никогда не бывает равнинных участков.
Первая фаза соответствует накоплению GI и GT. Она является результатом фотосинтетической функции листьев и завершается для основания побега до листопада. Она не совсем совпадает с фазой вызревания, но включает ее в себя.
Эта фаза начинается очень рано (молодые междоузлия уже богаты крахмалом) и продолжается, развиваясь прогрессивно, что особенно четко выражено у сорта Уньи белый. Иными словами, на графиках, касающихся однолетних побегов, начало отражало бы значительное изменение способности побега к росту. Наоборот, начало вызревания двухлетних побегов отмечено очень четко внезапным ростом содержания GI, причем этот рост наступает после фазы его уменьшения, начавшейся в период распускания.
Вторая фаза характеризуется уменьшением GI и GT, что соответствует гидролизу части накопившихся веществ. Примечательно то, что фаза гидролиза GI наступает непосредственно вслед за фазой синтеза. Между ними не существует никакого перехода, и количество GI или GT не стабилизируется ни разу в течение годичного цикла. Впрочем, количество GT не задерживается на уровне, достигнутом во время листопада. Оно уменьшается так же, как количество GI, но в значительно меньшей пропорции. Это уменьшение легко объяснимо: в основном оно соответствует потреблению углеводов, необходимых для дыхания побегов. Что же касается GI, как будет видно ниже, вопрос сложнее.
Третья фаза характеризуется новым ростом содержания GI и GT. Она совершенно отлична от первой в том смысле, что не зависит от тех же причин, ибо протекает после листопада.
Кроме того, следует объяснить и особый вопрос: в зависимости от сорта и условий года бывает так, что к моменту распускания почек количество GI в побегах оказывается большим, чем во время листопада (в конце октября), и во всех случаях содержание GT, достигнув минимума (в конце второй фазы), вновь возрастает.
Практические выводы
Невозможно определить с точностью степень вызревания побега в период покоя, основываясь лишь на содержании в нем GI. Действительно, сразу видно, что один и тот же побег, представляющий собой один и тот же морфологический вид в течение всего периода покоя, можно считать вполне или недостаточно вызревшим в зависимости от момента, когда он был срезан с куста. Так, например, если взять два крайних случая, можно считать его не вполне вызревшим в январе и вполне вызревшим в апреле, причем разница в содержании GI в эти два срока может достигнуть 10 и более процентов.
Ввиду того, что крахмал входит в состав большей части GI, тест при помощи йода не слишком подходящ для определения качества побегов, так как при его применении содержание этого полисахарида должно оставаться практически постоянным после листопада. Из рис. 1 явствует, что содержание GT — лучший критерий, но он требует двух определений — GI и GST.
Использование теста с йодом в практике виноградарства является иллюстрацией ошибочного приравнивания содержания крахмала к содержанию общих запасов. Как известно, тест, предложенный в 1901 г. Ravazn Bonnеt, позволяет, по мнению этих авторов, уточнить степень вызревания побегов и даже распределить их в четыре категории: слабо, средне, хорошо и очень хорошо вызревшие.
СПАДИЛО.РУ
Теория для подготовки к блоку №3 ОГЭ по биологии: система, многообразие и эволюция живой природы.
Теория для подготовки к блоку №4 ЕГЭ по биологии: система и многообразие органического мира.
Корень
Корень – подземный вегетативный орган высших растений, обладающий неограниченным ростом в длину.
Функции корня
- Закрепление растения в субстрате
- Всасывание, проведение воды и минеральных веществ
- Запас питательных веществ
- Взаимодействие с корнями других растений, грибами, микроорганизмами, обитающими в почве (микориза, клубеньки бобовых)
- Вегетативное размножение
- Синтез биологически активных веществ
- У многих растений корни выполняют особые функции (воздушные корни, корни-присоски)
- Видоизменения и специализация корней
- Корни некоторых строений имеют склонность к метаморфозу
Корни бывают различны, а именно могут видоизменяться.
Видоизменения корней
- Корнеплод — видоизменённый сочный корень. В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
- Корневые клубни – корнеклубни (корневые шишки) образуются в результате утолщения боковых и придаточных корней (тюльпаны, георгины, картофель).
- Воздушные корни — боковые корни, растут вниз. Поглощают дождевую воду и кислород из воздуха. Образуются у многих тропических растений в условиях повышенной влажности.
- Микориза — сожительство корней высших растений с гифами грибов. При таком взаимовыгодном сожительстве, называемом симбиозом, растение получает от гриба воду с растворёнными в ней питательными веществами, а гриб — органические вещества. Микориза характерна для корней многих высших растений, особенно древесных. Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.
- Бактериальные клубеньки на корнях высших растений — сожительство высших растений с азотфиксирующими бактериями — представляют собой видоизменённые боковые корни, приспособленные к симбиозу с бактериями. Бактерии проникают через корневые волоски внутрь молодых корней и вызывают у них образование клубеньков.
- Дыхательные корни — у тропических растений — выполняют функцию дополнительного дыхания.

- главный корень
- боковые корни
- придаточные корни
Главный корень развивается из зародышевого корешка. Боковые корни возникают на любом корне в качестве бокового ответвления. Придаточные корни образованы побегом и его частями.
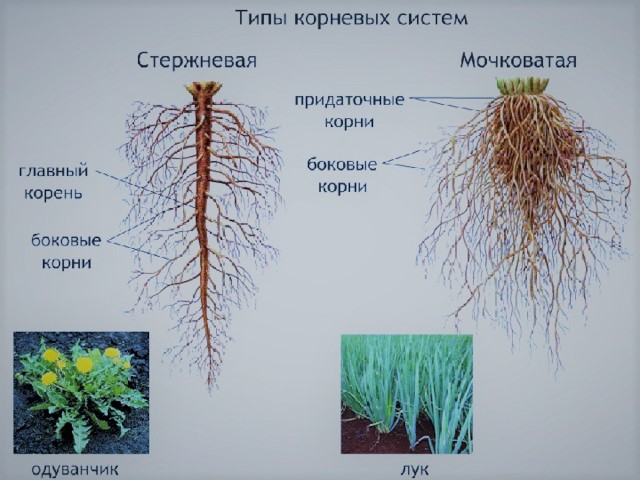
Совокупность корней одного растения называют корневой системой.
Виды корневых систем
- Стержневая
- Мочковатая
- Ветвистая
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди других корней (характерна для двудольных). Стержневая корневая система проникает в почву обычно глубже, чем мочковатая.
В мочковатой корневой системе на ранних этапах развития главный корень, образованный зародышевым корешком, отмирает, а корневая система составляется придаточными корнями (характерна для однодольных). Мочковатая корневая система лучше оплетает прилегающие частицы грунта, особенно в его верхнем плодородном слое.
В ветвистой корневой системе преобладают одинаково развитые главный и несколько боковых корней (у древесных пород, земляника).
Побег
Побег – это стебель с расположенными на нем листьями и почками.
Составными частями побега являются стебель, листья, почки. При прорастании семени из зародышевой почечки формируется первый побег растения — его главный побег, или побег первого порядка. Из главного побега формируются боковые побеги, или побеги второго порядка, а при повторении ветвления — третьего порядка и т. д. Придаточные побеги формируются из придаточных почек.
Так формируется система побегов, представленная главным побегом и боковыми побегами второго и последующего порядков. Система побегов увеличивает общую площадь соприкосновения растения с воздушной средой.
Побег, на котором образуются цветки, называется цветоносным побегом, или цветоносом (иногда термин «цветонос» понимают в более узком смысле — как участок стебля, на котором находятся цветки).
Вегетативный невидоизменённый побег — единый орган растения, состоящий из стебля, листьев и почек, формирующийся из общего массива меристемы (конуса нарастания побега) и обладающие единой проводящей системой. Стебли и листья, являющиеся основными структурными элементами побега — часто рассматриваются как его составные органы, то есть органы второго порядка. Кроме того, обязательная принадлежность побега — почки. Главная внешняя черта, отличающая побег от корня — наличие листьев.
В сезонном климате умеренных широт рост и развитие побегов из почек носит периодический характер. У кустарников и деревьев, а также у большинства многолетних трав это происходит один раз в году — весной или в начале лета, после чего формируются зимующие почки будущего года, а в конце лета — осенью рост побегов заканчивается.
Строение побега
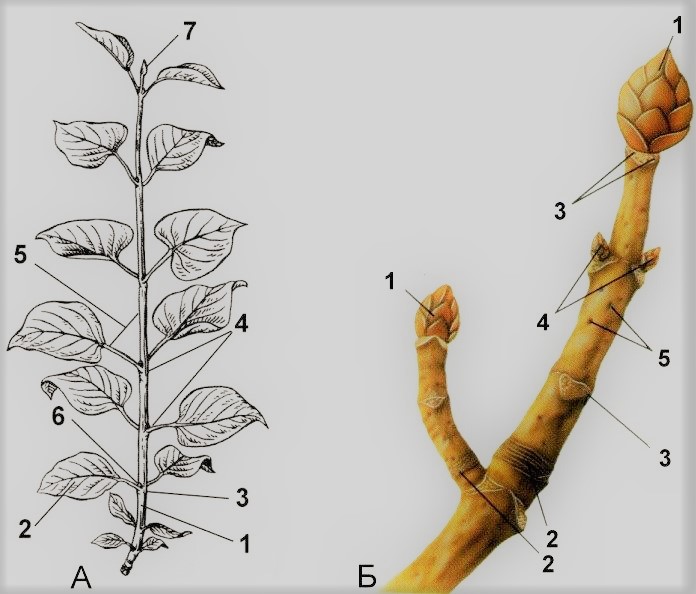
А (с листьями). 1 – стебель; 2 – лист; 3 – узел; 4 – междоузлие; 5 – пазуха листа; 6 – пазушная почка; 7 – верхушечная почка.
Б (после листопада). 1 – верхушечные почка; 2 – почечные кольца; 3 – листовые рубцы; 4 – боковые почки.
Типы побегов
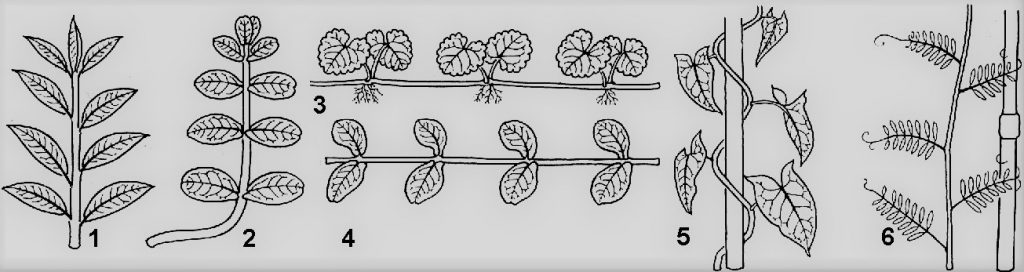
1 – прямостоячий; 2 – приподнимающийся; 3 – ползучий; 4 – стелющийся; 5 – вьющийся; 6 – лазающий.
Видоизменения побегов
- Колючка — сильно одревесневающий безлистный укороченный побег с острой верхушкой. Колючки побегового происхождения выполняют главным образом защитную функцию. У дикой яблони, дикой груши, крушины слабительной (Rhamnus cathartica) в колючки превращаются укороченные побеги, имеющие ограниченный рост и оканчивающиеся остриём.
- Усик — жгутовидный ветвистый или неразветвлённый побег метамерного строения, в типичном случае лишённый листьев. Стеблевые усики, как узкоспециализированный побег, выполняют опорную функцию.
- Корневище — подземный побег с чешуевидными листьями низовой формации, почками и придаточными корнями. Толстые, сильно разветвлённые ползучие корневища характерны для пырея, короткие и довольно мясистые — для купены, ириса, очень толстые — для кубышки, кувшинки.
- Стеблевой клубень — видоизменённый побег с ярко выраженной запасающей функцией стебля, наличием чешуевидных листьев, которые быстро сшелушиваются, и почек, формирующихся в пазухах листьев и называемых глазками (топинамбур).
- Луковица — подземный (реже надземный) сильно укороченный специализированный побег, в котором запасные вещества откладываются в чешуях листовой природы, а стебель преобразован в донце. Луковица — типичный орган вегетативного возобновления и размножения. Луковицы свойственны однодольным растениям из семейства Лилейные (лилия, тюльпан, лук), Амариллисовые (амариллис, нарцисс, гиацинт) и др. Как исключение, они встречаются и у двудольных — у некоторых видов кислицы и жирянки.
- Клубнелуковица — видоизменённый подземный укороченный побег с толстым стеблем, запасающим ассимилянты, придаточными корнями, отрастающими с нижней стороны клубнелуковицы, и сохраняющимися засохшими основаниями листьев (плёнчатые чешуи), в совокупности составляющими защитный покров. Клубнелуковицы имеют безвременник, гладиолус, иксия, шафран.
Стебель
Стебель — удлинённый побег высших растений, служащий механической осью, также выполняет роль производящей и опорной базы для листьев, почек, цветков.
Классификация стеблей
По расположению относительно уровня почвы:
По степени одревесневания:
- травянистые
- деревянистые (например, ствол — главный многолетний стебель дерева; стебли кустарников называют стволиками)
По направлению и характеру роста:
- прямостоячие (например, подсолнечник)
- лежачие (стелющиеся) — стебли лежат на поверхности почвы, не укореняясь (вербейник монетчатый)
- приподнимающиеся (восходящие) — нижняя часть стебля лежит на поверхности почвы, а верхняя поднимается вертикально (сабельник)
- ползучие — стебли стелются по земле и укореняются благодаря образованию в узлах придаточных корней (будра плющевидная)
- цепляющиеся (лазящие) — прикрепляются к опоре с помощью усиков (горох)
- вьющиеся — тонкие стебли, обвивающие опору (луносемянник)
По форме поперечного сечения:
- округлые
- сплюснутые
- трёх—, четырёх—, многогранные (гранистые)
- ребристые
- бороздчатые (желобчатые)
- крылатые — стебли, у которых по острым граням тянутся плоские травянистые выросты (чина лесная) или низбегающие на стебель основания листьев (окопник лекарственный)
Строение стебля
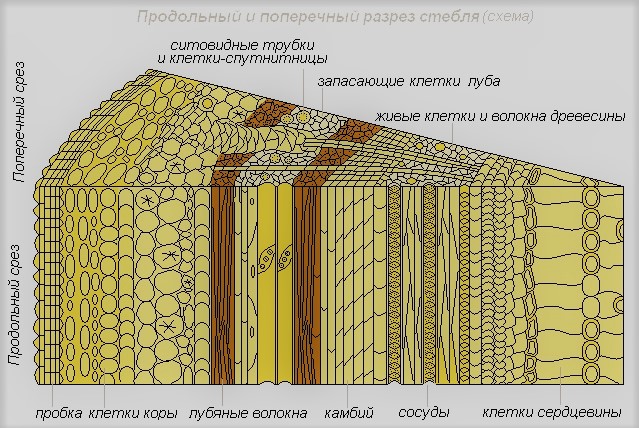
Снаружи стебель защищен покровными тканями. У молодых стеблей весной клетки покровной ткани покрыты тонкой кожицей. У многолетних растений к концу первого года жизни кожица замещается многослойной пробкой, состоящей из мертвых клеток, заполненных воздухом. Для дыхания в кожице (у молодых побегов) имеются устьица, а позже образуются чечевички – крупные, рыхло расположенные клетки с большими межклетниками.
К покровной ткани прилегает кора, образованная разными тканями. Наружная часть коры представлена слоями клеток механической ткани с утолщенными оболочками и тонкостенных клеток основной ткани. Внутренняя часть коры образована клетками проводящей ткани и называется лубом.
В состав луба входят ситовидные трубки, по которым идет нисходящий ток: органические вещества передвигаются от листьев. Ситовидные трубки состоят из клеток, соединенных концами в длинную трубку. Между соседними клетками имеются мелкие отверстия. Через них, как через сито, передвигаются органические вещества, образующиеся в листьях.
Ситовидные трубки остаются живыми недолго, чаще 2-3 года, изредка – 10-15 лет. На смену им постоянно образуются новые. Ситовидные трубки составляют небольшую часть в лубе и обычно собраны в пучки. Кроме этих пучков в лубе имеются клетки механической ткани, главным образом в виде лубяных волокон, и клетки основной ткани.
К центру от луба в стебле расположена другая проводящая ткань – древесина.
Древесина образована разными по форме и величине клетками и состоит из сосудов (трахей), трахеид и древесных волокон. По ним идет восходящий ток: вода с растворенными в ней веществами передвигается от корней к листьям.
В центре стебля лежит толстый слой рыхлых клеток основной ткани, в которых откладываются запасы питательных веществ, – это сердцевина.
У некоторых растений (георгина, тюльпан, огурец, бамбук) сердцевина занята воздушной полостью.
Между древесиной и лубом у двудольных растений находится тонкий слой клеток образовательной ткани –камбий. В результате деления клеток камбия увеличивается (растет) толщина стебля. Клетки камбия делятся вдоль своей оси. Одна из дочерних появляющихся клеток отходит к древесине, а другая – к лубу. Прирост особенно заметен в древесине. Деление клеток камбия зависит от сезонного ритма – весной и летом происходит активно (образуются крупные клетки), осенью замедляется (образуются мелкие клетки), а зимой останавливается. В итоге образуется годичный прирост древесины, хорошо заметный у многих деревьев, называемый годичным кольцом. По числу годичных колец можно подсчитать возраст побега и дерева в целом.
Ширина годичных колец у древесных растений зависит от условий окружающей среды. Так, в холодном климате, на болотных почвах величина годичных колец древесины очень мала. В благоприятных климатических условиях, на богатых почвах толщина годичных колец увеличивается. Сопоставляя чередование широких и узких годичных колец у ствола, можно определить, в каких условиях жило растение, а также установить колебания погодных условий за многие годы.
Функции стебля
- проводящая (главная функция)
Стебель служит опорой растению, он держит на себе тяжесть находящихся на нем листьев, цветков и плодов.
В стебле могут откладываться запасные питательные вещества. В этом проявляется запасающая функция стебля.С помощью стебля побег выносит свои листья и почки к свету в ходе роста растения. В этом проявляются важная осевая функция стебля и функция роста.
Лист — один из важнейших органов растений, основными функциями которого является фотосинтез, газообмен и транспирация.
Внутреннее строение листа
Лист состоит из следующих тканей:
- Эпидермис — слой клеток, которые защищают от вредного воздействия среды и излишнего испарения воды. Часто поверх эпидермиса лист покрыт защитным слоем восковидного происхождения (кутикулой).
- Паренхима — внутренняя хлорофиллоносная ткань, выполняющая основную функцию — фотосинтез.
- Сеть жилок, образованных проводящими пучками, состоящими из сосудов и ситовидных трубок, для перемещения воды, растворённых солей, сахаров и механических элементов.
- Устьица — специальные комплексы клеток, расположенные, в основном, на нижней поверхности листьев; через них происходит испарение воды и газообмен.
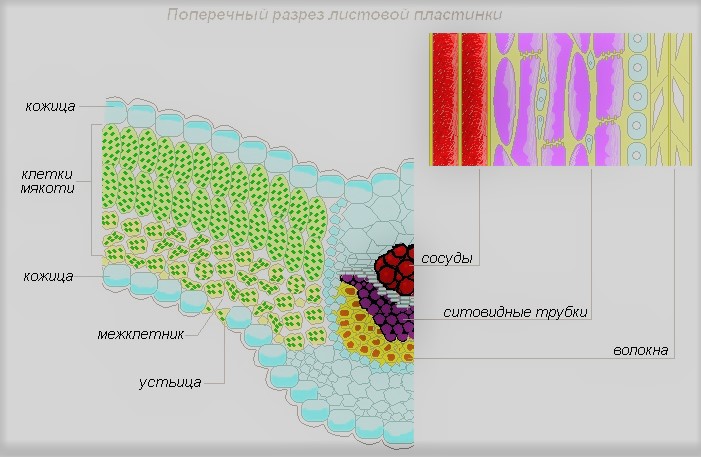
Внешнее строение листа
Лист внешне состоит из:
- черешка (стебелька листа)
- листовой пластинки (лопасти)
- прилистников (парных придатков, расположенных по обеим сторонам основания черешка)
- место, где черешок примыкает к стеблю, называется влагалищем листа
- угол, образованный листом (черешком листа) и вышерасположенным междоузлием стебля, называется пазухой листа
- в пазухе листа может образоваться почка (которая в этом случае называется пазушной почкой), цветок (называется пазушным цветком), соцветие (называется пазушным соцветием)
Не все растения имеют все части листьев, у некоторых видов парные прилистники чётко не выражены либо отсутствуют; может отсутствовать черешок, а структура листа может не быть пластинчатой.
Стеблевые части – Основы вызревания побегов винограда
Содержание материала
АНАТОМИЧЕСКИЕ, ФИЗИОЛОГИЧЕСКИЕ И БИОХИМИЧЕСКИЕ ОСНОВЫ ВЫЗРЕВАНИЯ ПОБЕГОВ
Вызревание побега виноградной лозы имеет большое практическое значение — с ним связана зимостойкость побегов и почек, а также качество привоя и подвоя при производстве посадочного материала. Поэтому проблема вызревания побега вызывает значительный интерес, особенно после нашествия филлоксеры.
Общие сведения
Исследованиями Gouin и Andouard (1899) установлено, что во время вызревания сухой вес побегов увеличивается, а содержание воды уменьшается. Снижается также удельный вес черенков. Далее эти же авторы (Gouin, Andouard, 1901) определяли содержание азота и крахмала в побегах, однако ясных выводов о роли этих веществ в процессе вызревания побегов не было сделано.
Kdvessi (1901) изучал процессы одревеснения, толщину клеточных мембран склеренхимы, содержание крахмала и величину крахмальных зерен. По его определению, хорошее вызревание побега обусловлено развитием перидермы, дифференцировкой тканей и величиной крахмальных зерен. Rаvaz и Bonnet (1901), однако, считают, что размер клеточных оболочек не имеет значения для зрелости побега. Как эти авторы, так и ряд других исследователей (Schmitthenner, 1909) не установили разницы в толщине клеточных оболочек хорошо и плохо вызревших побегов.
Исследования Ravaz и Bonnet по вызреванию побега виноградной лозы охватывали ряд показателей (цвет коры, диаметр сердцевины, отношение сердцевина—древесина, удельный вес, содержание органических и неорганических веществ и др.), однако положительная корреляция установлена лишь между степенью вызревания и содержанием СаО и крахмала.
Gasраr (1904) определял диаметр вызревших побегов, удельный вес, содержание воды, сухого вещества и золы, эфирную и спиртную вытяжки, содержание крахмала и др. Эти исследования также не дают полного представления о физиологии и биохимии вызревания.
Schmitthenner (1909) придает наибольшее значение перидермальной оболочке — между состоянием зрелости и образованием перидермы он устанавливает прямую зависимость. По его мнению, наиболее важной функцией перидермы является защита побега от лишнего испарения воды зимой. Хорошо вызревшие побеги зимой выделяют значительно меньше воды, чем плохо вызревшие, поэтому величину транспирации можно использовать в качестве показателя зрелости побегов. Schmitthenner считает также, что для процесса вызревания побегов имеет значение степень дифференциации флоэмы и ксилемы и содержание в них питательных веществ, в особенности крахмала. Между накоплением крахмала и закладкой перидермы имеется прямая связь. Гемицеллюлозу он считает конституционным элементом, поскольку им не установлено динамики содержания пентозанов в побегах.
В своих исследованиях Bernatsky (1916) обратил внимание на толщину побегов, состояние диафрагмы, толщину и дифференциацию флоэмы. Kroemer (1923) же придает большое значение при вызревании побегов процессам до закладки феллогена.
Мержаниан (1948) изучал динамику вызревания побега виноградной лозы (V. vinifera) на основании наблюдений за изменением окраски коры. Он установил, что вызревание побега первых 3—7-и междоузлий протекает медленно, в течение 10—15 дней. Далее наступает очень интенсивное вызревание побега — 8—14-го междоузлий в течение трех дней.
Аналогическое явление при вызревании побегов установлено также Димитровым (1962) при изучении некоторых агротехнических приемов выращивания подвоя Берландиери х Рипария Кобера 5 ББ.
Неодновременное вызревание побега установлено также Рябчуном (1965). Он показал, что процесс лигнификации побегов винограда начинается уже после образования первых элементов протоксилемы в нижних междоузлиях и распространяется с ростом стебля в длину и толщину, по его оси в акропетальном направлении, а по радиусу — в центробежном (для древесины) и центростремительном (для коры) направлениях.
Мержаниан (1951) рассматривает вызревание побегов как процесс анатомической и физиологической перестройки тканей, наступающей после закладки феллогена. Наиболее важные анатомические изменения, наблюдающиеся в процессе вызревания побегов, это образование корковой ткани и утолщение клеточных стенок в результате накопления в них лигнина, гемицеллюлозы и минеральных веществ. В числе важнейших физиологических изменений Мержаниан отмечает уменьшение содержания воды в клетках и накопление крахмала и других защитных веществ.
Изучению анатомической и биохимической стороны процесса вызревания побегов виноградной лозы посвящено исследование Орлова (1958). Однако наиболее обстоятельное изучение биологии вызревания виноградного побега, как нам представляется, проведено аспирантом Петровичем (1955). Поэтому мы отметим наиболее важные результаты этого исследования*, оставшиеся до сих пор неопубликованными.
*Исследование проводилось под руководством проф. К. Д. Стоева на побегах подвойного сорта Берландиери х Рипария Кобера 5 ББ и включало изучение ряда анатомических, морфологических и биохимических показателей.
Источники:
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/steblevye-chasti-14.html
http://spadilo.ru/koren-pobeg-stebel-list/
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/steblevye-chasti-2.html





