
Химия и энергетика дыхания – Современные представления о природе молекулярных механизмов и путях энергообмена зеленого растения
Химия и энергетика дыхания – Современные представления о природе молекулярных механизмов и путях энергообмена зеленого растения
Химизм и энергетика дыхания;
Процесс дыхания состоит из двух фаз: анаэробной, происхо-дящей в цитоплазме клеток, и аэробной, осуществляемой в ми-тохондриях.
Необходимо отметить тесную связь между процессами дыхания и брожения, на которую впервые обратил внимание С. П. Костычев. Первый этап этих процессов аналогичен и заключается в бескисло-родном распаде глюкозы на две молекулы пировиноградной кисло-ты (ПВК). Он получил название гликолиза. Дыхание и все виды бро-жения связаны между собой через пировиноградную кислоту:
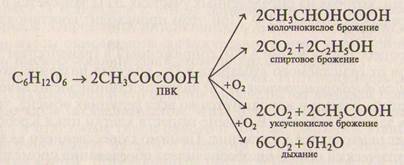
Гликолиз состоит из ряда реакций, каждая из которых катали-зируется специфическим ферментом.
Пировиноградная кислота поступает в митохондрии, где осу-ществляется вторая — аэробная фаза дыхания. Далее она окисля-
ется до СО, и Н,О в цикле Кребса и электронно-транспортной цепи, при этом высвобождается энергия в виде АТФ. В результате окисления одной молекулы глюкозы образуется 38 молекул АТФ.
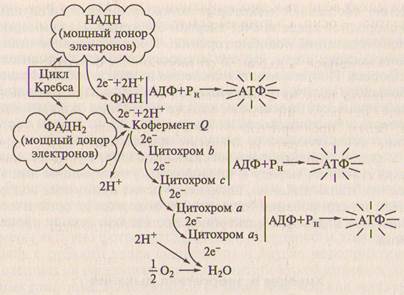
Рис. 9.3. Передача энергии по цепи переносчиков электронов в мито-хондриальной мембране.
Процесс высвобождения энергии заключается в передаче элект-ронов от богатых энергией соединений — доноров (НАДН и ФАДН ) по электронно-транспортной цепи (ЭТЦ) на активиро-ванный кислород (рис. 9.3). Этот поток электронов можно срав-нить с каскадом водопадов. Переносчики электронов — ферменты в ЭТЦ — расположены по мере убывания восстановленности, по-этому транспорт электронов сопровождается высвобождением энергии, которая на определенных участках ЭТЦ запасается в мак-роэргических связях АТФ. При этом происходит восстановление 1/2 О2 до Н 2О.
Процесс образования АТФ, сопряженный в транспортом элек-тронов от окисляемого субстрата к О3, получил название окисли-тельного фосфорилирования. АТФ является единой энергетической «валютой» клетки и используется во всех реакциях обмена, требу-ющих затраты энергии. В обмене веществ клетки цикл Кребса за-нимает центральное положение. Помимо существенного вклада в энергетический обмен он обеспечивает образование промежуточ-ных соединений, которые используются в синтезе различных ве-ществ в клетке.
Дыхание представляет собой процесс, противоположный фо-тосинтезу. Эти процессы тесно связаны друг с другом. Углеводы, полученные при фотосинтезе за счет солнечной энергии, при дыхании окисляются и выделяют эту энергию, которая поддер-живает жизнь растения, животных, в том числе и человека. Един-ство и противоположность процессов ассимиляции — создания органических веществ и диссимиляции — распада их с выделением энергии — основа жизни на Земле.
Химия и энергетика дыхания – Современные представления о природе молекулярных механизмов и путях энергообмена зеленого растения
Знаменитая теория дыхания Палладина, кратко сформулированная в виде двух уравнений, которая произвела полный переворот в обычном понимании дыхательного процесса и теперь вошла в основные руководства по физиологии растений запечатлена в следующих двух уравнениях:
С6Н12О6 + 12R + 6Н2О = 6СО2 + 12RН2,
12RН2 + 12R + 12Н2O.
Основным уравнением, выражающим собой принципиальную сущность дыхательного процесса, является первое. В этой первой фазе дыхательный материал (глюкоза) уже полностью распадается до своих простейших элементов – СО2 и водорода, но водород не выделяется в свободном состоянии, а захватывается при помощи редуктаз (дегидраз) водородными акцепторами, без которых распад глюкозы вообще не может иметь места. (процесс идет чисто анаэробно). Кислород к этому основному акту дыхания не имеет никакого отношения. Весь СО2 – анаэробного происхождения, и уносимый с нею кислород заимствуется не из внешней атмосферы, а изнутри системы. Дыхание для своего осуществления не нуждается ни в кислороде, ни в оксидазах. Основными ферментами в этой фазе дыхания являются дегидразы.
Кислород воздуха вмешивается в акт дыхания лишь во второй фазе, но, принципиально рассуждая, играет здесь второстепенную роль. Он должен восстановить активность водородных акцепторов, освободив их от захваченного водорода. Кислород окисляет не углерод дыхательного субстрата, а уже отщепленный от него водород, перенесенный на акцепторы. Сам кислород выступает лишь в роли последнего заключительного водородного акцептора.
Эта принципиально новая точка зрения на процесс дыхания, выраженная Палладиным в столь ясно отчетливой форме, не была еще при нем достаточно широко обоснована экспериментальными данными и явилась в значительной мере плодом научной интуиции автора, хотя в ней вместе с тем мы должны видеть, как бы подведение итогов многолетней работы Палладина над процессом дыхания.
Одновременно с Палладиным проблемой дыхания занимались в целом ряде крупнейших научно-исследовательских институтов и лабораторий Западной Европы. Наибольшую популярность приобрели две новые школы – Виланда и Варбурга.
Т.Виланд развивал взгляды на роль дегидраз и водородных акцепторов, вполне аналогичные взглядам Палладина. Расхождение их теорий заключалось в том, что Виланд категорически отрицал какую бы то ни было роль оксидаз как специфических активаторов кислорода, считая молекулярный кислород способным самостоятельно отнимать водород от водородного акцептора. По мнению же Палладина, водородные акцепторы не могут самопроизвольно освобождаться от водорода, но требуют для этого участия оксидаз, которые поэтому являются обязательным фактором в реакции, выраженной во втором уравнении Палладина.
Противник Виланда, Варбург, считал, что молекулярный кислород не может вступить в организме в какой бы то ни было окислительный процесс, если в организме отсутствует система железоорганических соединений, типичным представителем которых он считал геминфермент. Варбург утверждал, что геминфермент активирует молекулярный кислород, т.е. как бы дает первый толчок к началу окислительных процессов, и без него никакой дыхательный процесс не может совершаться. Далее, по мнению Варбурга, окислительный импульс через промежуточные звенья (геминовые соединения) доходит до дыхательного субстрата и окисляет его. Варбург утверждал, что дыхание осуществляется путем активации кислорода, а отнюдь не водорода. Но ведь Палладин как раз и говорил о той же необходимости активации молекулярного кислорода, защищая перед Виландом роль оксидаз в процессе дыхания.
Все различие в основных посылках Варбурга и Палладина заключается в том, что первый, работая по преимуществу с объектами животного происхождения, называл свой активатор молекулярного кислорода геминферментом, а Палладин, работавший с объектами растительного происхождения, сохранил за этим активатором ранее установившееся в науке название оксидазы. Но по существу оба говорили об одном и том же, протестуя против непримиримой позиции Виланда, отрицавшего необходимость энзиматической активации молекулярного кислорода.
Согласно взглядам большинства современных научных авторитетов Запада обе теории – и Виланда, и Варбурга – оказываются правильными, они охватывают различные фазы одного и того же чрезвычайно сложного процесса. Современная наука находит разрешение вопроса в синтезе обеих теорий, называя современное учение о внутреннем механизме дыхательных процессов теорией Виланда–Варбурга.
Современные представления о природе молекулярных механизмов и путях энергообмена зеленого растения – Биохимическая стадия фотосинтеза
Содержание материала
Биохимическая (темповая) стадия фотосинтеза
Вопрос о путях превращения неорганического углерода СO2 в углерод органических соединений и регулирующих эти процессы механизмах служил и продолжает служить предметом изучения на протяжении многих десятков лет.
Неоценимую роль в раскрытии закономерностей превращения ассимилируемой СO2 сыграли исследования, в которых использовались прогрессивные современные методы молекулярной биологии (меченые атомы, различные виды хроматографии и радиоавтографии, электрофорез, гемофильтрация, скоростное центрифугирование и др.).
С помощью этих методов достоверно доказано, что в подавляющем большинстве случаев первым стабильным продуктом фотосинтеза, образующимся за счет ассимилируемой углекислоты, является фосфоглицериновая кислота (ФГК), имеющая формулу
СН2O(Р) — CHOH—CHOH
Удалось также выяснить пути образования ФГК. Она возникает в результате присоединения СO2 к молекуле пятичленного сахара — рибулезодифосфата (РДФ) и последующих превращений образующихся при этом промежуточных соединений. Присоединение СO2 к РДФ приводит к образованию двух молекул ФГК, которая восстанавливается за счет энергии мобилизуемых пигментами электронов и превращается в фосфоглицериновый альдегид (ФГА). Именно на этом этапе и осуществляется акт превращения неорганического углерода в органический. Далее следует ряд сложных взаимных превращений углеводов с различной длиной углеродной цепи. В ходе этих превращений имеет место образование молекулы рибулезофосфата. Для полной регенерации молекулы рибулезодифосфата, выполняющего, как указывалось выше, роль первичного акцептора СO2, используется макроэргическая фосфатная связь АТФ. Синтез последней, как известно, осуществляется в ходе фотофосфорилирования. Регенерация молекулы РДФ — обязательное условие, от которого зависит возможность непрерывного протекания процесса фотосинтеза.
Следовательно, в цикле синтеза сахаров имеются 3 взаимосвязанные фазы: 1) вторичное фосфорилирование рибулезофосфата, его превращение в рибулезодифосфат, являющийся акцептором СO2; присоединив СO2, РДФ делится на две частицы ФГК; 2) восстановление ФГК в ФГА; 3) регенерация акцептора СO2 — РДФ.
Таков один из генеральных путей ассимиляции СO2. В определенных условиях акцептором электронов, образующихся в результате фотодиссоциации воды, может служить ион NO3, восстановление которого приводит к образованию в ходе фотосинтеза аминокислот. Установлено также, что характер превращений ФГК зависит от специфических особенностей обмена веществ отдельных видов растений, условий их корневого питания, интенсивности и спектрального состава света и т. п. В ряде случаев первым этапом превращений ФГК является ее вторичное фосфорилирование. Роль первичного акцептора СO2 в определенных условиях выполняет фосфоэнолпировиноградная кислота (ФЭП).
В начале 60-х годов было установлено, что у довольно обширной группы растений первичное карбоксилирование ФЭП является доминирующим путем ассимиляции СO2. К этим растениям относятся кукуруза, сахарный тростник, сорго, просо, злаковые травы и др. Таким образом был открыт по существу новый путь ассимиляции СO2, получивший название С-4, в отличие от вышеописанного, известного под именем С-3.
Для растений группы С-4 характерна очень высокая интенсивность фотосинтеза, которая, в свою очередь, обусловлена особенностями анатомического строения их листьев. Так, у этих растений обнаружены специализированные хлоропласта, которыми заполнены клетки паренхимы обкладок, окружающие имеющуюся в листьях густую сеть мелких проводящих пучков. Хлоропласты обкладки имеют более крупные размеры при сравнении с хлоропластами клеток мезофилла. Весьма важна их способность накапливать большие количества крахмала. Для этих хлоропластов характерно, кроме того, наличие сильно развитого периферического ретикулума, хорошо выраженная ламеллярная структура и отсутствие и, во всяком случае, слабая развитость гран.
Преобладание C-4-пути ассимиляции СO2 не исключает, а наоборот, предполагает постоянное его взаимодействие с С-3-путем. Такое кооперирование может достигаться посредством переброски части СO2, поглощенной ФЭП на РДФ, иными словами, посредством ее включения в цикл С-3. Этим взаимодействием, сосредоточением карбоксилирующих ферментов в тех и других ассимилирующих клетках обеспечивается быстрое перемещение продуктов ассимиляции из клеток мезофилла в клетки обкладок, а из них — в проводящие органы. Таковы особенности организации фотосинтетического аппарата растений группы С-4, обуславливающие характерную для них высокую интенсивность процессов роста и большую продуктивность.
Заканчивая обзор современного состояния знаний о природе фотосинтеза, следует еще раз подчеркнуть, что эта кардинальная функция биоэнергетики представляет собой сложную совокупность строго координированных, организованных в пространстве и во времени окислительно-восстановительных реакций. Уникальный характер этой функции определяется ее направленностью против градиента, следствием чего является увеличение количества свободной энергии в системе. Источником этого возрастания является энергия фотонов, абсорбируемых специфической группой соединений — пигментами.
Отличительной особенностью последних является то, что поглощаемая шли световая энергия не деградирует и не рассеивается в мировом пространстве, а преобразуется в энергию химическую. Наряду со светозависимыми реакциями, фотосинтез включает большое число реакций темновых, осуществляющихся биохимическим путем. Первичной фотохимической реакцией фотосинтеза является фотоокисление реакционного центра — пигмента, входящего в состав ФСБ. При окислении пигмента происходит разделение зарядов, в результате которого образуется свободный, богатый энергией электрон, а также вакансия (дырка). Эта реакция протекает с очень большой скоростью
(10-11—10-12s). Последовательная передача электронов по цепи промежуточных переносчиков сопряжена с процессами реализации освобождающейся на этом пути энергии, на базе которой и возникает АТФ. С участием этих аккумуляторов энергии осуществляется заключительный этап фотосинтеза — восстановление неорганического углерода СO2 и образование стабильных органических соединений (углеводов, белков, жиров и др.). Помимо последних, непосредственно в ходе процессов фотосинтеза образуются различные физиологически активные соединения, в числе которых пигменты, специфические нуклеиновые кислоты (РНК и ДНК), специфические белки, включая большое число различных белков — ферментов и др. Все эти соединения являются участниками процесса фотосинтеза. Составляющее основу фотосинтеза сочетание световых и темновых реакций, их увязка в единое целое, синхронность множества составляющих эту функцию элементарных реакций, противоположных по своей физико-химической природе, их координация и объединение в общий, целенаправленный поток обусловлены специфической структурно-морфологической организацией органоидов, в которых сосредоточена данная функция. Особая роль принадлежит в этом отношении ярко выраженной гетерогенности структуры мембран и стромы, включающих ламеллы, граны, тилакоиды, квантосомы и др. Весьма гетерогенным является и химический состав хлоропластов. Так, сюда входят комплексы пигментов, различающихся по химическим, физическим и функциональным свойствам, специфические белки и нуклеиновые кислоты, липиды, обширный набор различных ферментов, витамины и многие другие соединения.
Осуществляющийся в специализированных органоидах-хлоропластах фотосинтез, вместе с тем, отнюдь не является функцией локальной, изолированной от всех других функций клетки и организма в целом. Ход процессов фотосинтеза самым тесным образом связан со всем комплексом процессов жизнедеятельности. Характер этой связи непостоянен, он находится в зависимости от влияния самых различных условий, включая генетически обусловленные свойства растения и условия, в которых осуществляется его развитие. С этим именно и связаны широкие возможности управления фотосинтезом в интересах практики.
Источники:
http://studopedia.su/15_120770_himizm-i-energetika-dihaniya.html
http://vseobiology.ru/fiziologiya-rastenij/1648-22-sovremennye-predstavleniya-o-khimizme-dykhaniya
http://vinograd.info/knigi/fiziologiya-vinograda-stoev/sovremennye-predstavleniya-o-prirode-molekulyarnyh-mehanizmov-i-putyah-energoobmena-zelenogo-rasteniya-3.html




