
Динамика нарастания листа – Лист
Динамика нарастания листа – Лист
Динамика нарастания листа – Лист
Морфогенез побега
Основные части побега – стебель, листья, почки, цветки и др. – закладываются в апикальной меристеме побега, которая является производной эмбриональной ткани дистального конца зародыша.
Апекс побега. Апекс (конус нарастания, точка роста) вегетирующего побега семенного растения состоит из меристематических клеток, которые по своему размеру, частоте и направленности делений, по особенностям метаболизма могут быть разделены на несколько зон, прежде всего на тунику и корпус. Туника, или мантия, – это один, два или более слоев клеток, покрывающих апекс снаружи. Клетки туники делятся преимущественно антиклинально ( т.е. плоскость деления перпендикулярна поверхности апекса). Из наружного слоя туники образуется эпидермис. Все остальные клетки, лежащие под туникой, входят в состав корпуса, в котором при более дробной анатомо-физиологическом разделении апикальной меристемы побега на зоны выделяют центральную, периферическую и стержневую меристемы. Дистальная группа клеток в тунике и центральной (аксиальная) зона выполняют функцию инициалей. Клетки этих участков апекса относительно крупные и делятся сравнительно редко. Периферическая зона (инициальное кольцо) состоит из мелких меристематических клеток, которые интенсивно делятся. В них число рибосом выше чем у инициальных клеток. Клетки этой зоны формируют примордии (зачатки) боковых органов побега – листьев и почек. Граница между туникой и корпусом в этой зоне исчезает. Клетки все зон апекса имеют крупные ядра, густую цитоплазму и не содержит вакуолей.
Стержневая (колончатая) зона состоит из вакуолизированных клеток с относительно низким содержанием РНК. Клетки этой зоны делятся главным образом антиклинально и дают начало продольным рядам клеток первичной коры и сердцевины стебля. Границы между описанными зонами в апексе побега весьма условны и далеко не всегда различимы.
Конус нарастания побега, обладая высокой способностью к саморазвитию, тем не менее, нуждается в притоке не только питательных веществ, но и фитогормонов. Изолированные апексы с двумя-тремя листовыми примордиями нормально развиваются лишь в том случаи, если в питательной инкубационной среде присутствуют цитокинин, а в ряде случаев – и ауксине.
Рост и развитие листа
Формирующийся лист проходит четыре фазы: 1) образование примордия; 2) формирование оси листа; 3) закладка пластинки листа за счет боковой меристемы; 4) рост пластинки растяжением.
Каждый листовой примордий образуется в виде бугорка в периферической меристеме апекса побега благодаря локальным периклинальным делением клеток (плоскость деления параллельна поверхности апекса). У многих видов периклинальные деления в зоне закладки примордия происходит и в тунике. Примордий пазушной почки появляется несколько позже. В нем затем формируется апикальная меристема, гомологичная апексу главного побега.
Период времени между заложением двух листовых зачатков называется пластохроном. Продолжительность его у разных видов и даже у одного и того же вида при разных условиях сильно варьирует: от нескольких часов до нескольких суток. Примордии листьев образуются на апексе в строго заданной последовательности, предопределяя расположение листьев на зрелом побеге, или филлотаксис. У растений распространен спиральный филлотаксис. Замечено, что на апексах с многочисленными примордиями угол между ними близок к значению 137,5о . при таком угле в идеальном случае ни один лист на стебле не располагается точно под другим, что обеспечивает их минимальную затененность. По теории В. Гофмейстера, такое листорасположение достигается путем возникновения новых листовых зачатков в промежутках между уже существующими примордиями ( «теория доступного пространства»).
Согласно «теории отталкивания», предложенной Ю.Шоуте, когда детерминируется центр листового примордия, в нем продуцируется специфические вещества, ингибирующие образование новых центров в непосредственной близости от заложившегося. Соответственно новый примордий развивается вне ингибиторных полей соседей.
Эти гипотезы довольно хорошо согласуются друг с другом, так как «доступное пространство» может быть обусловлено не только поверхностной зоной между примыкающими примордиями, но и зоной, от их ингибирующего влияния. В этом месте и закладывается новая примордия. Возникшие зачатки листьев оказывают влияние на нижележащие ткани, индуцируя дифференциацию проводящих пучков. Это действие обусловлено делением ауксина, который синтезируется в формирующихся примордиях.
Верхушечные клетки конуса листового примордия делятся особенно интенсивно, превращая бугорок в пальцеобразный выступ. Этот выступ состоит в основном из клеток будущей средней жилки и черешка листа. На краях зоны средней жилки начинает функционировать маргинальная (краевая) меристема, дающая начало листовой пластинке. Одновременно с этим прекращается верхушечный рост листа.
Инициальные клетки маргинальной меристемы и клетки самой этой меристемы делятся главным образом антиклинально, что приводит к увеличению листовой пластинки, а не ее толщины. Поверхностные инициальные маргинальные клетки образуют эпидермис, а субмаргинальные инициальные клетки – внутренние ткани листа.
После 8-9 циклов делений клетки маргинальной меристемы переходят к растяжению. Эпидермальные клетки кончают делиться первыми, но продолжают расти растяжением. Клетки губчатой паренхимы прекращают делиться растут раньше других тканей. Поэтому продолжающееся деление и растяжение эпидермиса приводят к тому, что губчатые клетки отодвигаются друг от друга, образуя большие межклетники. Палисадные клетки делятся и растут со скоростью, близкой к росту эпидермиса. Этот процесс останавливается несколько раньше окончания растяжения эпидермиса. Поэтому и палисадные клетки не сколько отрываются друг от друга, образуя небольшие межклетники.
Особенность роста листа однодольных растений состоит в том, что деления в возникшем на одной стороне апекса листовом бугорке распространяются в обе стороны и охватывают всю окружность стебля. Возникающий серповидный (у злаков) или кольцевой (у осок) меристематический валик дает начало листу, растущему вверх. До формирования валика бугорок растет, как и у двудольных, верхушкой. Листовая пластинка удлиняется путем интеркалярного роста, который более продолжителен у основания пластинки.
На рост листьев большое влияние оказывают периодичность, качество и интенсивность света. Свет сине-фиолетовой части спектра тормозит рост междоузлий и способствует росту листьев ( у двудольных). Интенсивное освещение способствует развитию палисадной ткани.
Гормональная регуляция роста листа изучена недостаточно. Показано, что цитокинин и ауксин необходимы для формирования и развития примордиев и тканей листа, ауксин принимает участие в образовании жилок, гиббереллин способствует более интенсивному росту листовой пластинки в длину.
Рост листьев ограничен в отличии от пазушных почек, у которых заложенные апексы будут функционировать длительное время, если эти почки дадут боковые побеги.
Динамика нарастания листа – Лист
Лист – вегетативный орган растения, располагающийся на побеге. Место расположения листа на побеге называется узел. Узел (лат. nodus) — участок побега (стебля) растения, от которого отходят боковые органы (ветви, листья, почки, придаточные корни и другие.)
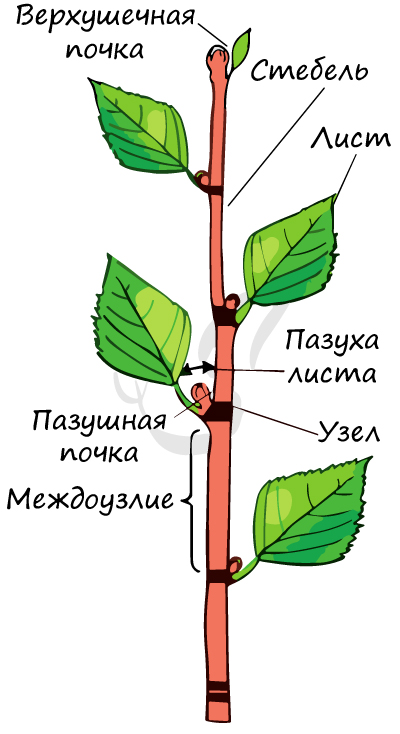
Строение и функции листа
Основная ткань пластинки листа – мезофилл. Выделяют столбчатый и губчатый мезофилл, функции которых различны.
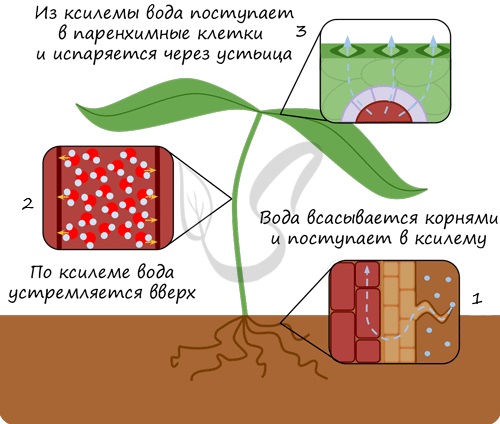
Благодаря наличию хлоропластов в клетках столбчатой ткани мякоти листа происходит процесс фотосинтеза, в результате которого образуется большое количество органических веществ, доставляемых флоэмой в разные части растения. Вообразите следующую информацию в виде 3D-модели: проводящая система листа является продолжением проводящей системы стебля, в месте узла происходит отхождение сосудисто-волокнистого пучка в направлении листа.
В губчатой ткани листа расположены межклетники, вход в которые открывают устьица. Здесь происходит газообмен между организмом растения и внешней средой, заключающийся в процессах дыхания и фотосинтеза. Крайне важно разделить два понятия: фотосинтез и дыхание.
Не удивляйтесь тому, что растения поглощают кислород в процессе дыхания. Все живые клетки аэробных организмов находятся в процессе дыхания постоянно, днем и ночью. Запомните, что дыхание это поглощение кислорода и выделение углекислого газа. В ходе светозависимой фазы фотосинтеза напротив, кислород выделяется как ненужный побочный продукт, а углекислый газ поглощается клетками.

Осуществляется через устьица в эпидермисе (кожице).
Самые первые листья растения – зародышевые листья (семядоли или семенодоли), которые развиваются у зародыша ещё в семени. В дальнейшем листья формируются из примордиев – нерасчленённых зачатков листьев в виде бугорков или валиков на конусе нарастания побега.

Основные части листа
- Основание листа
Это расширенная нижняя часть листа. У некоторых растений оно, разрастаясь, преобразуется в незамкнутую или замкнутую трубку, которую называют листовое влагалище.
Выполняет главные функции листа – газообмен и фотосинтез, в основании пластинка сужается и переходит в стеблевидный черешок.
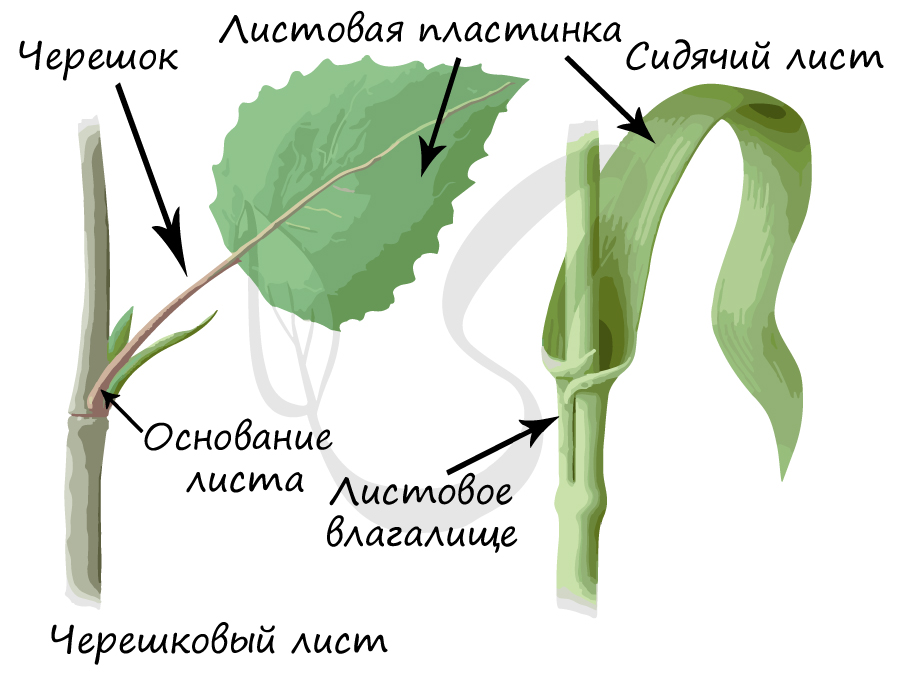
Это тонкая стеблевидная часть листа, идущая от листовой пластинки к узлу побега.
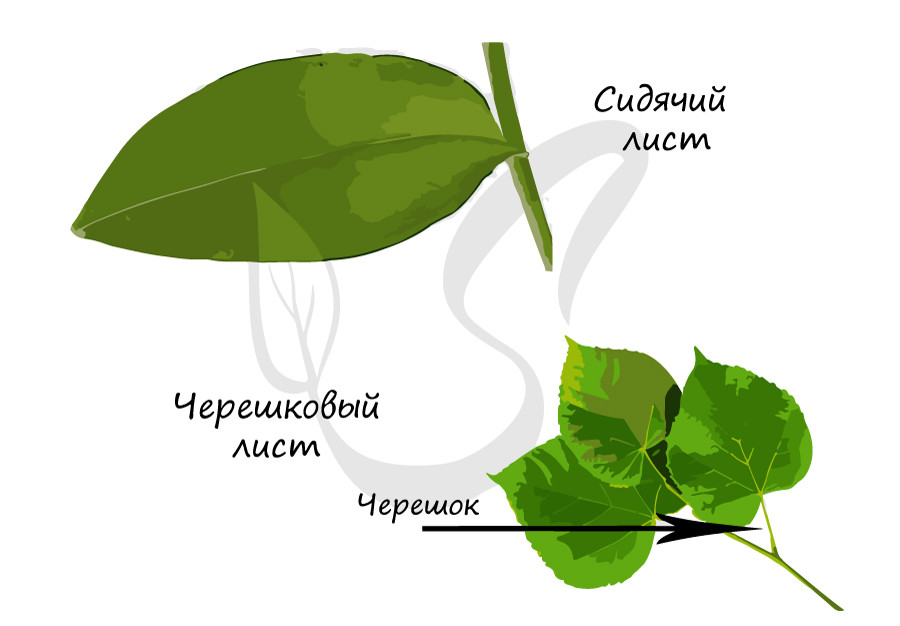
Меняя свою форму, черешок смещает листовую пластинку. Таким образом, основная функция черешка – обеспечить как можно большую освещенность листовой пластинки, вынести листья к свету. Именно так и создается листовая мозаика – расположение листьев на растении, при котором они не затеняют друг друга. Листья с черешками называются черешковыми, без черешка – сидячими.
Выросты листообразной формы, расположенные у основания листа. Они могут срастаться со стеблем или быть свободно расположенными. У многих растений прилистники отсутствуют в принципе, или образуются, но рано отмирают.
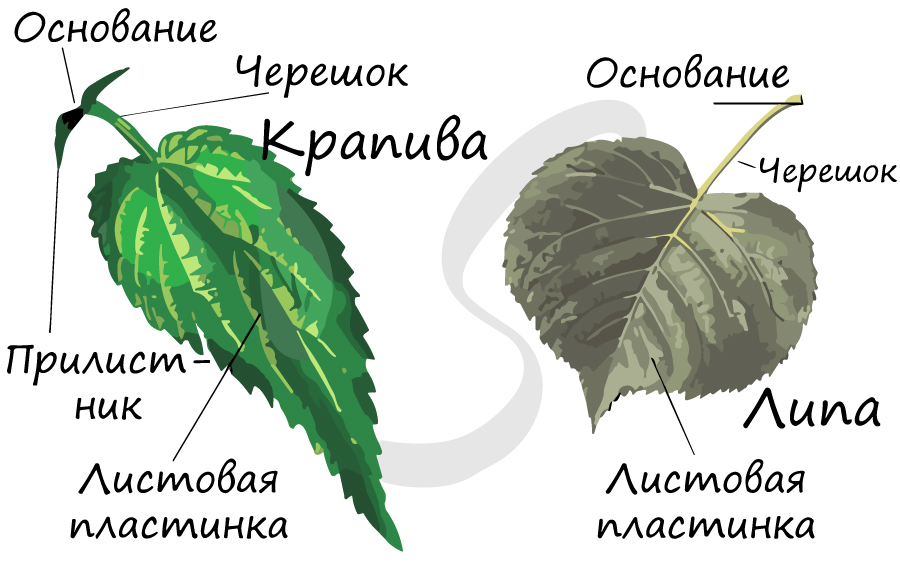
Лист называют полным, если в составе его элементов имеется пластинка, основание, прилистники и черешок. Полные листья характерны для многих широко известных растений: рябина, дуб, черемуха, роза.
Лист называют неполным, если у него отсутствует черешок (сидячий лист), прилистники или пластинка. Сидячие листы, лишенные прилистников, имеют лен, гвоздика, алоэ. Отсутствуют прилистники также на листьях картофеля, сирени, капусты. В редких случаях лист может не иметь листовой пластинки, тогда ее функции перенимают черешок – у акации, прилистники – у чины безлисточковой.

Жилкование листьев
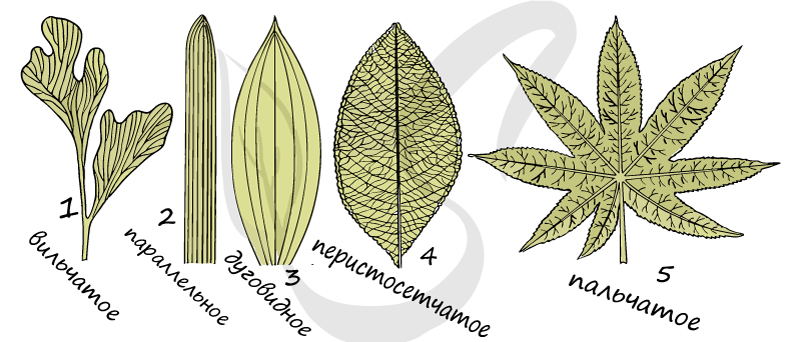
Это обозначение системы жилок с проводящими пучками в листовой пластинке. Жилкование листьев бывает:
-
Вильчатое (дихотомическое) жилкование
Встречается у многих папоротниковидных растений и примитивных семенных, при вильчатом жилковании жилки делятся дихотомически (одна жилка разделяется на две жилки).
При таком типе жилкования крупные жилки проходят вдоль листовой пластинки параллельно друг другу. Характерно для злаковых растений.
Отличается наличием крупных жилок, которые подобно дуге изогнуты вдоль листовой пластинки. Характерно для однодольных.
Перистое (перисто-сетчатое) жилкование
Для этого типа характерна выраженная центральная (главная) жилка, от которой в стороны отходят более тонкие боковые ветви. Имеется у дуба черешчатого, черемухи обыкновенной.
Пальчатое (пальчато-сетчатое жилкование)
Такой тип жилкования отличается наличием нескольких примерно одинаковых по размеру крупных жилок, расходящихся веером по пластинке, при этом сходящихся в одной точке у ее основания. Имеется у манжетки обыкновенной, клена платановидного.
Форма листа
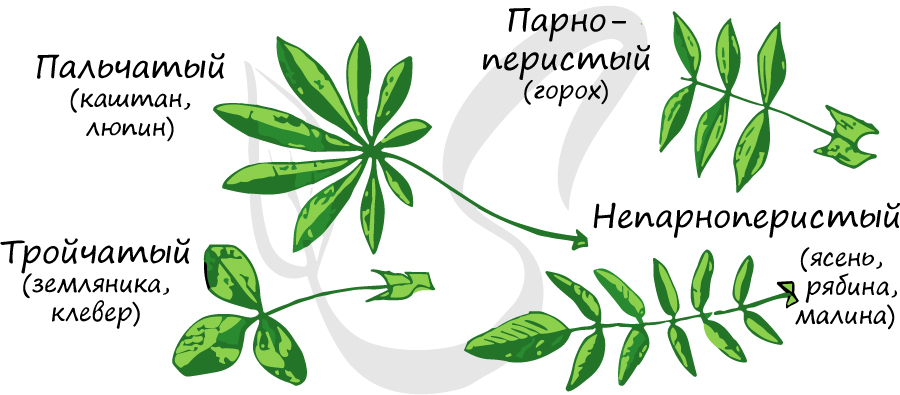
Листья подразделяются на простые и сложные. Лист называют сложным, если несколько листовых пластинок, прикрепленных собственными короткими черешочками, располагаются на одном общем черешке (рахисе). Каждую листовую пластинку сложного листа называют листочком или пластиночкой. Сложные листья (названия которых говорят сами за себя) бывают:
- Однолисточковые – у мандарина, лимона.
- Тройчатосложные – у земляники, клевера.
- Пальчатосложные, состоящие из множества листовых пластинок, у люпина, каштана конского.
Необходимо особо отметить, что есть сложные листья, у которых листочки расположены по всей длине рахиса – пункты 4 и 5.
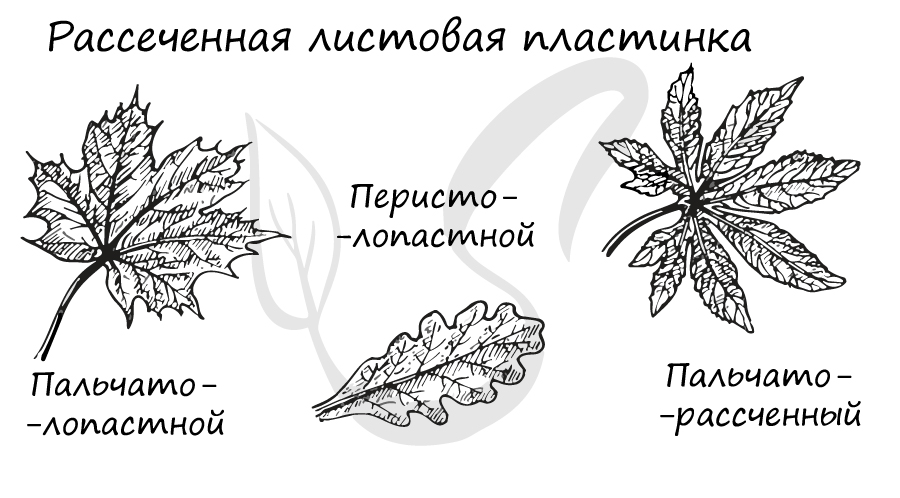
Простым листом называется лист, состоящий из одной листовой пластинки. Простые листья подразделяются на несколько типов, в зависимости от структуры листовой пластинки. Простые листья могут быть:
- С цельной листовой пластинкой – сирень, береза, тополь, яблоня.
- С рассеченной (расчлененной) листовой пластинкой. Каждую отдельную часть простой пластинки называют сегментом. Здесь также имеется еще одно деление, по степени расчлененности листовых пластинок различают:
- Пальчтолопастную (перилопастную) – в случае если расчленение не превышает 1/3 всей поверхности листовой пластинки.
- Перистолопастную (перистораздельную) – расчленение не превышает половины (до 1/2) листовой пластинки.
- Пальчаторассеченную (перисторассеченную) – расчленение достигает главной жилки листа или основания листовой пластинки.
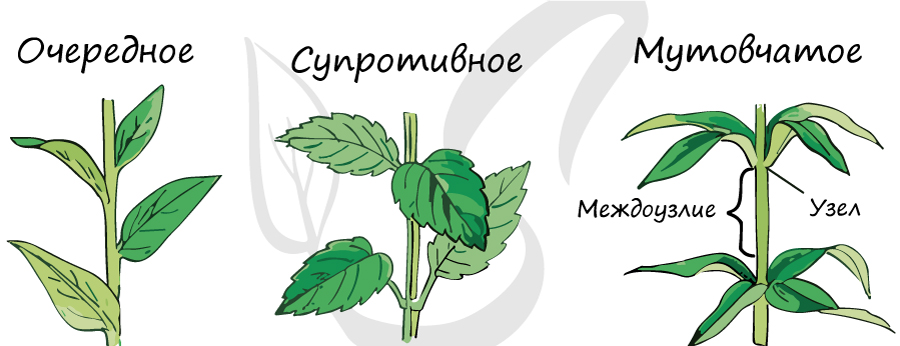
Листорасположение
Представляет собой порядок расположения листьев на стебле. Выделяют следующие типы листорасположения:
- Очередное – от узла отходит только один лист. Имеется у березы, липы, дуба.
- Супротивное – на узле располагаются два листа, супротив (напротив) друг друга. Встречается у бузины, клена, калины.
- Мутовчатое – на узле стебля 3 и более листьев. Есть у вороньего глаза, ветренницы, элодеи.
Видоизменения листьев
Это интереснейшие приспособления, которые возникли в процессе приспособления растений к различным средам обитания. Они выполняют дополнительные функции, но главная их задача – это адаптация растения к условия среды.
Не все растения питаются автотрофно, для некоторых из них свойственен гетеротрофный тип питания. Известный пример росянка капская – насекомоядное растение. Ее лист покрыт липкой вязкой массой, которая выделяется волосками листа. Если насекомое садится на лист, то приклеивается к нему, волоски начинают сворачиваться, и насекомое оказывается в образовавшейся полости. После чего начинается выделение ферментов в полость и переваривание насекомого.

Образования, которые выполняют опорную функцию. Цепляясь усиками за опору, растение занимает в пространстве вертикальное положение, растут вверх. Имеются у чины, гороха.

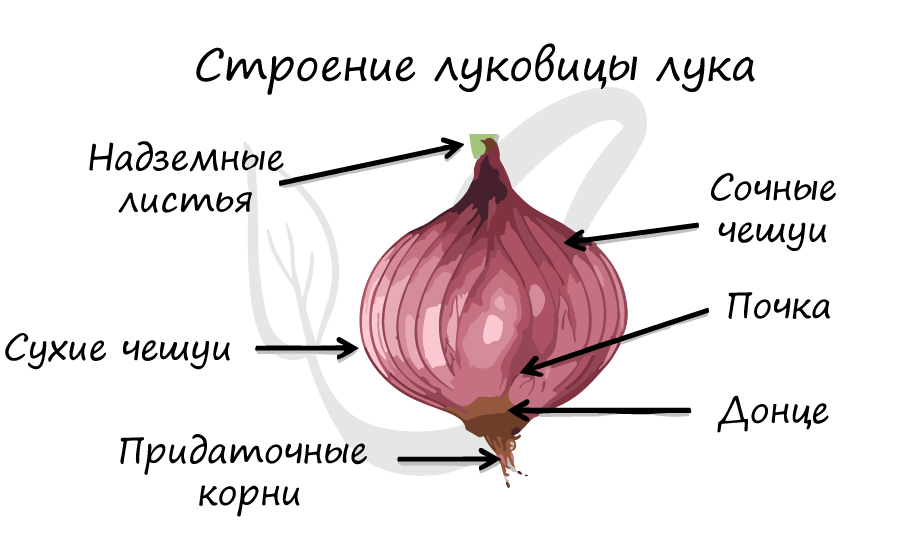
Выполняют различные функции. К примеру, чешуи почки защищают ее от механических повреждений, а листья у лука в луковице превращены в сочные чешуи, которые запасают питательные вещества.
Ограждают растение от поедания его животными. Подобную защитную функцию выполняют колючки барбариса, кактуса.

Эти видоизменения листьев не утратили свою основную функцию, и приобрели дополнительную – запасание воды. Особенно актуальна эта функция для растений суккулентов, произрастающих в местах с засушливым климатом – алоэ, молодил, очиток.

Хвоя – это видоизмененные листья голосеменных (хвойных) растений. Таким листьям, в отличие от неизмененных, нужно меньше питательных веществ и воды. Они способны противостоять холоду и засухе, при этом выполняя свою основную роль – обеспечение процесса фотосинтеза.
Вечнозелеными растениями является подавляющее большинство голосеменных, которые сохраняют хвою в течение 12 месяцев, не сбрасывая ее перед зимой. У отдельных видов, сосны долговечной, хвоя сохраняется до 45 лет.

©Беллевич Юрий Сергеевич
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Развитие листа. Длительность жизни листьев. Листопад, его механизм и значение. Метаморфозы листьев.
В своем развитии лист проходит две фазы: внутрипочечную и внепочечную. В течение первой фазы листовой зачаток увеличивается главным образом за счет деления клеток. При этом он постепенно приобретает форму, характерную для взрослого листа. Однако в почке лист остается миниатюрным, сложенным или свернутым. При переходе во вторую фазу лист развертывается. Во второй фазе он сильно разрастается за счет деления и растяжения клеток. Листовой зачаток, не имеющий признаков дифференциации, называют примордием. Вначале он растет равномерно за счет деления клеток во всех направлениях. Но вскоре рост его дифференцируется, становится неравномерным. Обычно раньше всего прекращают деления клетки верхушки листового зачатка. После этого он может расти только за счет интеркалярной и краевой меристемы. Довольно рано зачаточный лист дифференцируется на 2 части: базальную (нижнюю) и апикальную (верхнюю). Развитие этих частей дальше идет неодинаково. Из базальной части развивается основание листа (а также прилистники, влагалище – если они есть), из апикальной части – пластинка и черешок.
Достигнув окончательных размеров, лист может жить в течение разного времени, однако по сравнению с осевыми органами, листья многолетних растений недолговечны. У большинства растений они живут несколько месяцев, а у вечнозеленых растений от 1,5 до 20 лет. Вечнозелёность этих растений объясняется тем, что старые листья постепенно заменяются новыми, т. е. у них нет единовременного опадения всех листьев.
Наибольшей продолжительностью жизни отличаются листья хвойных. Так, у сосны обыкновенной лист живёт 2-4 года, а у ели — 5-7 лет, тиса — 6-10 лет. У одних и тех же видов растений при подъёме в горы и с продвижением на север длительность жизни листьев увеличивается. Так, у ели обыкновенной в Хибинах хвоя живёт 12-18 лет.
ЛИСТОПАД опадение листьев обычно у деревьев и кустарников, реже у трав (крапива, недотрога). Листья могут опадать одновременно все в определённый период года (напр., у листопадных деревьев) или постепенно по одному в течение длительного времени (у вечнозелёных растений). Листопадные деревья во влажнотропич. лесах стоят без листьев иногда всего неск. дней, в умеренном поясе — до 8—9 мес. Л. – нормальный физиол. процесс, связанный со старением листьев. Перед Л. в листьях происходят глубокие биохимич., физиол. и структурные изменения. Хлорофилл обычно разрушается, каротиноиды сохраняются дольше и обусловливают осеннюю окраску листьев. Питат. вещества из листьев оттекают в запасающие органы (клубни, корневища и т. п.) к точкам роста, к растущим молодым листьям. Механизм Л. связан с появлениему основания листа (или осн. черешка) отделительного слоя из легко разъединяющихся паренхимных клеток. Проводящие пучки, удерживающие лист на стебле, разрываются под тяжестью листа и порывами ветра. Л. — выработанное в процессе эволюции приспособление к уменьшению в не благоприятных условиях поверхности наземных органов, что сокращает потерю влаги и предотвращает поломку ветвей под тяжестью снега.
Метаморфозы листьев – выработанные в ходе эволюции необратимые изменения формы листьев в результате приспособления органов растения к условиям среды обитания (т. е. с выполнением листьями новых функций).
1. Колючки— одно из наиболее часто распространенных видоизменений; они служат защитой от поедания животными. При этом лист либо целиком превращается в колючку (кактусы, молочаи, барбарис, белая акация, верблюжья колючка), либо в колючку превращается его часть (бодяк, чертополох, падуб).
2. Усики(у сложных листьев некоторых видов растений) цепляются за опору, вынося весь побег к свету. При этом в усик могут превращаться либо верхние листочки сложного листа (горох, вика), либо весь лист целиком, а функцию фотосинтеза выполняют прилистники (некоторые виды чины).
3. Запасающую функцию выполняют сочные чешуилуковиц (лук, чеснок), листья алоэ, кочана капусты.
4. Кроющие чешуипочек защищают нежные зачаточные листья и конус нарастания от неблагоприятных условий внешней среды.
5. Ловчие аппаратыобеспечивают жизнь насекомоядных растений на болотах в условиях недостатка азота и других элементов минерального питания. Листья таких растений изменились до неузнаваемости, превратившись в ловушки (венерина мухоловка), кувшинчики (непентес). Листья некоторых растений своими блестящими, ярко окрашенными капельками на волосках привлекают муравьев, мух, комаров, других мелких насекомых; выделяющийся при этом сок содержит пищеварительные ферменты (росянка).
Анатомическое строение типичного листа.
Анатомическая структура листа формируется в конусе нарастания одновременно с формированием стебля. В начале формирования лист растёт своей верхней частью, затем верхушечный рост затухает, и зона роста сохраняется лишь у основания листа. У папоротников лист растёт верхушкой в течение всей жизни.
С обеих сторон лист покрыт эпидермой, защищающей внутренние ткани листа от неблагоприятных воздействий внешней среды. Газообмен и испарение воды осуществляется через устьица. Между двумя слоями эпидермы находится мезофилл, илихлоренхима, составляющий основную массу листа (рис. 37).

Рис. 37. Анатомическое строение листа камелии японской (Camellia japonica): 1 — верхняя эпидерма; 2 — столбчатый мезофилл; 3 — губчатый мезофилл; 4 — нижняя эпидерма; 5 — опорные клетки; 6 — собирательные клетки, 7 — собирательные клетки с друзами; 8 — устьице; 9 — ксилема; 10 — флоэма; 11 — склеренхима
Столбчатый мезофилл
В листьях, расположенных горизонтально, мезофилл дифференцирован на столбчатую и губчатую ткань. Столбчатый мезофилл примыкает к верхней стороне листа. Его клетки вытянуты перпендикулярно к поверхности листа, в них содержится много хлоропластов, основная функция его — фотосинтез. Наиболее хорошо выражена эта ткань у растений, произрастающих в условиях интенсивной освещённости. У тенелюбивых растений, наоборот, столбчатый мезофилл менее выражен, длина его клеток незначительно превышает их ширину.
Губчатый мезофилл
К нижнему эпидермису примыкает губчатый мезофилл, который состоит из округлых, рыхло расположенных клеток. Здесь имеются большие воздушные полости, которые сообщаются с устьицами. Основная функция этой ткани — газообмен и испарение воды, хотя фотосинтез в её клетках также происходит. У водных и болотных растений в мезофилле образуются крупные воздушные полости, превращающие его в аэренхиму.
У листьев, расположенных вертикально, нет резкого различия между столбчатой и губчатой тканью (у злаков). В условиях засушливого климата столбчатый мезофилл располагается и на нижней стороне листа (у лебеды).
Проводящие пучки
В мезофилле находятся проводящие пучки, образующие жилки. Чаще всего ониколлатеральные, причём ксилема в пучке повёрнута к верхней, а флоэма — к нижней стороне листа. Проводящие пучки листа обычно лишены камбия, т. е. являются закрытыми. Проводящие элементы листа отграничены от межклетников и клеток мезофилла плотно сомкнутыми обкладочными клетками. Крупные жилки содержат склеренхиму, а очень тоненькие жилки имеют упрощённое строение: ксилема может включать один-два проводящих элемента, а флоэма — одну ситовидную трубку с клеткой-спутницей.
77.243.189.108 © studopedia.ru Не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования. Есть нарушение авторского права? Напишите нам | Обратная связь.
Отключите adBlock!
и обновите страницу (F5)
очень нужноИсточники:
http://studbooks.net/916278/estestvoznanie/morfogenez_pobega
http://studarium.ru/article/8
http://studopedia.ru/20_17237_razvitie-lista-dlitelnost-zhizni-listev-listopad-ego-mehanizm-i-znachenie-metamorfozi-listev.html





