
Биосинтез липидов – Метаболизм липидов и их роль
Биосинтез липидов – Метаболизм липидов и их роль
Биосинтез липидов – Метаболизм липидов и их роль




Печень является главным местом синтеза жирных кислот, жиров, кетоновых тел и холестерина. Жиры могут также синтезироваться в жировой ткани, однако ее основной функцией остается депонирование липидов.
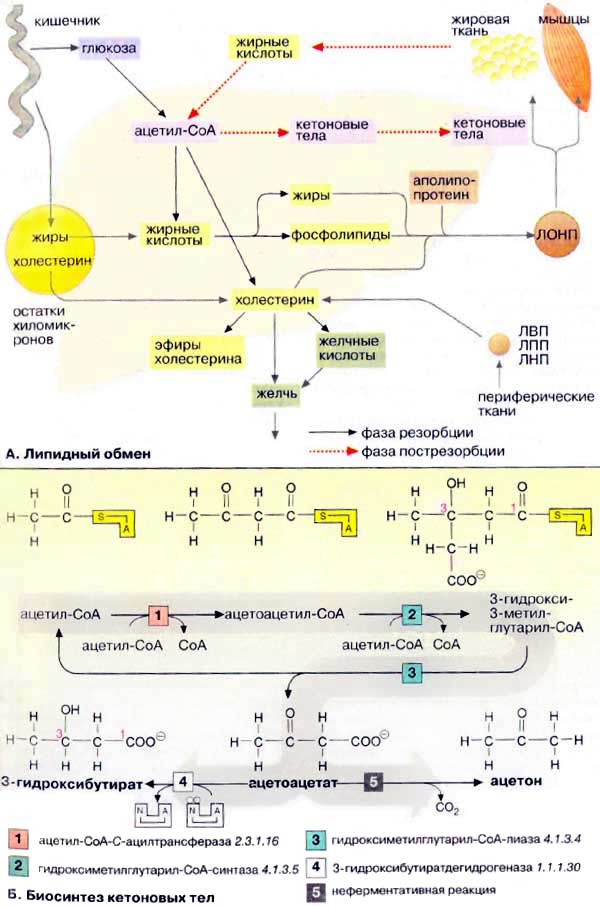
Обмен липидов в печени тесно связан с превращением углеводов и аминокислот. При поступлении питательных веществ в фазе резорбции (см. с. 300) глюкоза через промежуточное образование ацетил-КоА (ацетил-СоА) конвертируется в жирные кислоты. Печень может также извлекать жирные кислоты из липопротеинов, поступающих из желудочно-кишечного тракта (в виде хиломикронов) и других тканей (см. с. 272). Жирные кислоты используются для биосинтеза триглицеринов и фосфолипидов. При связывании жиров с аполипопротеинами образуются липопротеиновые комплексы очень низкой плотности [ЛОНП (VLDL), см. с. 272]. Они попадают в кровь и переносятся в другие ткани, прежде всего в жировую и мышечную ткань.
В фазе пострезорбции (см. с. 300), особенно в период поста или голодания, обмен липидов идет в обратном направлении, организм обращается к собственным запасам. В этих условиях жиры поступают из жировой ткани в кровь, переносятся в печень, распадаются в результате β-окисления до ацетил-КоА и, наконец, превращаются в кетоновые тела.
Холестерин поступает в организм из двух источников — с пищей и за счет эндогенного синтеза, причем большая часть холестерина синтезируется в печени. Биосинтез холестерина начинается с ацетил-КоА (см. с. 174). Полученный холестерин используется в синтезе желчных кислот (см. с. 306), встраивается в клеточные мембраны (см. с. 216), депонируется в жировых каплях в составе эфиров жирных кислот. Остальная часть поступает в кровь в составе липопротеиновых комплексов [ЛОНП (VLDL)] и переносится в другие ткани. Печень способствует обмену холестерина благодаря тому, что служит местом, худа поступают с кровью и где подвергаются расщеплению липопротеиновые комплексы [ЛВП, ЛПП, ЛНН (HDL, IDL, LDL), см. с. 272], содержащие холестерин и его эфиры с жирными кислотами.
При высокой концентрации ацетил-КоА в митохондриях гепатоцитов происходит конденсация двух молекул ацетил-КоА с образованием ацетоацетил-КоА [ 1 ]. Присоединение еще одной ацетильной группы [ 2 ] приводит к 3-гидрокси-З-метилглутарил-КоА (ГМГ-КоА) [ 3 ], который после отщепления ацетил-КоА превращается в ацетоуксусную кислоту (ацетоацетат) (цикл Линена). При восстановлении последней получается 3-гидроксибутират [ 4 ], а при неферментативном декарбоксилировании — ацетон [ 5 ]. Все три соединения принято называть «кетоновыми телами», что не совсем правильно, поскольку в 3-гидроксимасляной кислоте отсутствует кетогруппа!
Кетоновые тела поступают из печени в кровь, где они хорошо растворимы. Концентрация кетоновых тел в крови возрастает в фазе пострезорбции (фаза голодания). Наряду с жирными кислотами 3-гидроксибутират и ацетоацетат в этот период являются основными энергоносителями. Ацетон, не имеющий метаболической ценности, удаляется через легкие. После 1-2 недели голодания кетоновые тела начинают использоваться в качестве источника энергии нервными тканями. Однако при этом для обеспечения цитратного цикла необходимо минимальное количество глюкозы.
Если биосинтез кетоновых тел превышает потребности организма, они накапливаются в крови (кетонемия) и, наконец, выводятся с мочой (кетонурия). Оба феномена наблюдаются во время длительного голодания (углеводная недостаточность) и при заболеваний диабетом (Diabetes mellitis). Хотя 3-гидроксимасляная кислота является слабой кислотой (рКа примерно 4), возрастание концентрации кетоновых тел вызывает изменение рН в крови (кетоацидоз, см. с. 280). Кетонурия и кетоацидоз могут быстро привести к электролитному сдвигу (нарушению ионного гомеостаза) и потери сознания (кетоацидозной коме) и, следовательно, опасны для жизни.
Метаболизм липидов и их роль – Биосинтез липидов
Содержание материала
Основные пути метаболизма липидов микроорганизмами довольно хорошо изучены. Исследования, проведенные А. Гринбергом и опубликованные в 1968 г., представляют собой основную информацию об этих путях. Однако данные, касающиеся дрожжей, ограничены, поэтому мы приведем общий взгляд на пути синтеза и распада липидов в дрожжах.
Исследования, проведенные с помощью 1-С14, показали, что некоторые фракции митохондрий дрожжевых клеток способны синтезировать различные классы липидов. Именно в митохондриях и происходят основные процессы, например цикл трикарбоксильных кислот, окислительное фосфорилирование и цитохромные изменения.
Значительную роль в синтезе липидов играют и микросомы дрожжевой клетки.
Исследования Ф. Линена [132] показали, что образование жирных кислот осуществляется цитоплазматическим ферментным комплексом, а исходным субстратом при синтезе служит ацетил- КоА.
Лимитирующей стадией биосинтеза жирных кислот является ацетил
КоА-карбоксилазная реакция, в результате которой из ацетил-КоА при участии АТФ под действием фермента образуется малонил
КоА. Удлинение цепи жирных кислот при их биосинтезе происходит за счет приращения ацетильных остатков — малонил
S-АПБ (ацетилпереносящий белок) к карбоксильной группе ацетил
S-АПБ. Так, при образовании одной молекулы пальмитиновой кислоты взаимодействуют одна молекула активированного ацетила и семь молекул активированного малонила.
Как показали исследования Линена [132] и др., синтез жирных кислот с длинной цепью осуществляется дрожжами с помощью устойчивого мультиэнзимного комплекса (с молекулярной массой 2,3×106) в присутствии ацетил-коэнзима и малонил-КоА согласно реакции 
Дж. Ступе изолировал из высокоактивных дрожжей синтетазу жирных кислот в присутствии ингибитора протеаз. Ингибитор он применил для выделения нативного энзима, и этим исключил возможность деградации белковой молекулы энзима. С помощью электрофореза на полиакриламидном геле было показано, что выделенный энзим дает два пятна, молекулярная масса которых соответствует 212 000 и 203 000D.
М. В. Залешко и Г. А. Салохина [39] показали, что дрожжи как продуценты липидов в большей степени определяются составом жирных кислот. Присутствие в липидах дрожжей значительного количества ненасыщенных жирных кислот свидетельствует о том, что липиды дрожжей имеют сходство с растительными липидами, причем в них доминируют триглицериды (до 80%). Они содержат также моно- и диглицериды, фосфолипиды, стерины и их эфиры.
Образование липидов дрожжами в значительной степени зависит от дыхательной активности. Следует отметить, что интенсивность аэрации оказывает положительное влияние только до определенного предела, после чего излишняя аэрация даже вредно сказывается на синтезе липидов. Дрожжи, культивируемые при недостаточной аэрации среды, синтезируют значительное количество свободных жирных кислот, фосфолипидов и сравнительно мало образуют триглицеридов. При аэрации среды содержание ненасыщенных липидов возрастает.
Влияние температуры выращивания, pH среды и интенсивности аэрации существенно отражается на способности энзимов синтезировать липиды. По существу, эти факторы в известной степени управляют синтезом липидов.
Как показали исследования В. В. Залешко и Г. А. Салохиной [39], некоторые расы дрожжей способны образовывать наряду с интрацеллюлярными и экстрацеллюлярные липиды. Свойством
дрожжей синтезировать экзолипиды обладают почти все виды дрожжей. Синтез экзолипидов протекает в периплазматическом пространстве клеток, а затем эти липиды выделяются в окружающую среду.
Процесс синтеза липидов
Профилактика атеросклероза, как и терапия заболевания, напрямую связаны с контролем уровня липидных структур в организме. Особое внимание уделяется холестерину (ХС), молекула которого представляет собой липофильный спирт. Отсюда происходит непривычное на бытовом уровне, но химически корректное название вещества – холестерол. Именно окисление неиспользованных организмом липидов свободными радикалами – первый этап в последовательности формирования атеросклеротических бляшек. С другой стороны, соединения липидных структур с протеинами, создают биологические комплексы, способные очищать сосуды. Это липопротеины высокой плотности – ЛПВП. Таким образом, синтез и биосинтез липидов важен применительно к общему здоровью человека. Процесс непосредственно влияет на уровень холестерина в организме.

Что включает класс липидов
Категория объединяет жиры и подобные им вещества. На молекулярном уровне, липид формируется на двух базовых элементах: спирт и жирная кислота. Также допускается вхождение дополнительных компонентов. Подобные структуры относят к классу сложных липидов. Наибольший интерес, с точки зрения профилактики атеросклероза, привлекают следующие представители этого класса:
- Жирные спирты, а именно холестерин.
- Триглицериды.
Определенного внимания заслуживали бы жирные кислоты (ЖК), в частности полиненасыщенные – Омега-3. Вещество способствует снижению ХС. Однако организмом человека их синтез не осуществляется.
Общий принцип биосинтеза липидов
Образование ЖК и их производных начинается с цитоплазмы. Вторая часть биосинтеза – удлинение молекулярной цепи также продолжается в клетке, однако «производственная мастерская смещается» внутрь митохондрии. На каждом этапе, соединение обогащается двумя атомами C, что напоминает процесс бета-окисления, только в его обратной интерпретации.
Говоря более развернуто, в цитоплазме непосредственно и происходит синтез, например пальмитиновой кислоты. Митохондрии же, используют уже готовый «полуфабрикат», для производства полноценных жирных кислот, состоящих из 18-и и более атомов углерода. Выполнить весь биосинтез самостоятельно от «А» до «Я», митохондрии не в состоянии. Причина банальна – «низкий уровень квалификации». Возвращаясь к технической терминологии, митохондрии обладают очень низкой способностью включать меченые уксусные кислоты в длинную цепь липидных структур.

Хитрый трюк или как метаболит преодолевает митохондриальный барьер
Базовый внемитохондриальный биосинтез ЖК, напротив, не имеет общих пересечений с процессом их окисления. Его механизм, требует трех компонент:
- ацетил-КоА – первичный метаболит;
- CO2 – тут без комментариев, вещество общеизвестное;
- ионов бикарбоната – HCO3-.
Метаболит представляет собой строительный фундамент. Изначально ацетил-КоА образуется именно в митохондрии. Его синтез – следствие процесса окислительного декарбоксилирования. Просочиться напрямую в цитоплазму, соединение не может в силу непроницаемости для него митохондриальной мембраны. Удается осуществить проникновение путем обходного маневра:
- Митохондриальный метаболит производит цитрат, посредством взаимодействия с оксалоацетатом.
- Для синтезированного цитрата митохондриальная мембрана прозрачна. Поэтому его молекулы с легкостью пробиваются в цитоплазму.
- Далее происходит обратная трансформация. Едва преодолев мембрану, цитрат расщепляется на исходные компоненты – ацетил-КоА и оксалоацетат.
Таким образом, метаболит передается от митохондрии. В цитоплазме непосредственного получения соединения не происходит. Альтернативный вариант переноса ацетил-КоА возможен при участии карнитина. Однако, в процессе синтеза ЖК, – это своеобразный «бронепоезд, стоящий на запасном пути». Данный канал используется значительно реже.

Заключительный этап биосинтеза
Оказавшись в цитоплазме метаболит готов к производству прекурсора ЖК – малонил-КоА. Для этого ацетил-КоА и требуется двуокись углерода. Катализатором процесс выступает фермент ацетил-КоА-карбоксилазы. Биосинтез распределяется на два периода:
- Карбоксилирование биотин-энзима. Протекает в присутствии CO2 и АТФ.
- Перенос карбоксильной группы на метаболит.
Результирующий малонил-КоА в дальнейшем быстро трансформируется в ЖК. Процесс происходит с участием определенной ферментной системы. Фактически, это комплекс взаимосвязанных ферментов. Он именуется синтетаза жирных кислот, имеет 6 различных ферментов и связующий элемент – ацилпереносящий белок (осуществляют роль, аналогичную КоА).
Разобравшись с биосинтезом липидов на общем уровне, самое время перейти к конкретным примерам.
Биосинтез триглицеридов
Фундаментальными кирпичиками процесса выступают глицерин и ЖК. Изначально формируется промежуточный продукт – глицерол-3-фосфат. Это характерно для процессов биосинтеза, происходящих в почках и стенках кишечника. Клетки органов отличаются гиперактивностью фермента глицеролкиназы, чего нельзя сказать о мышечной и жировой ткани. Тут вещество формируется при помощи гликолиза – окисления глюкозы.
Эффект особенно проявляется при истощении. Вследствие этого, образуется недостаток глюкозы, что приводит к резкому снижению производства глицерол-3-фосфата. Как результат свободные ЖК не используются на ресинтез липидов, просто покидая ткань.
Еще одним источником промежуточного синтеза глицерол-3-фосфат выступает печень. Орган может производить вещество обоими способами, с использованием как глицерина, так и глюкозы, в качестве базового соединения. Впрочем, для синтеза глицерол-3-фосфат главное, что события произошли, поскольку в дальнейшем соединение ацилируется. То есть происходит процесс введения RCO- остатка при помощи молекул ЖК (КоА производные).
Это приводит к синтезу фосфатидной кислоты. В дальнейшем от полученного соединения отщепляется остаток фосфорной кислоты. Реакция дефосфорилирования протекает с участием фосфатидатфосфатазы. Ее продуктом выступает 1,2-диглицерид. До получения триглицерида остается единственный шаг. Он состоит в этерификации. Кислотой выступает 1,2-диглицерид, заместителем спирта – фермент ацил-КоА.

Биосинтез холестерина
Ферментативный процесс образования ХС – достаточно сложная «многоходовая комбинация», насчитывающая более 35 энзиматических реакций. Очевидно, что охватить подобный объем преобразований не под силу даже Остапу Бендеру. Поэтому проще рассмотреть базовые стадии биосинтеза холестерола:
- Получение мевалоновой кислоты. Происходит в эукариоте – домене живых организмов. Требует три молекулы активного ацетата.
- Формирование сквалена. Прекурсором выступает ранее произведенная мевалоновая кислота. Изначально соединение трансформируется в активный изопреноид, из 6 молекул которого и образуется сквален.
- Синтез холестерина. Процесс осуществляется циклизацией сквалена. Синтезируется своеобразный прекурсор – ланостерин, переход которого в ХС все еще находится под изучением.
Первоначально биосинтез инициируется формированием ацетоацетил-КоА. Далее, структура подвергается конденсации с 3-ей молекулой активного ацетата. Полученное производное вещество вступает в реакцию восстановления, что и приводит к формированию мевалоната.
Следующий шаг в биосинтезе ХС – превращение мевалоната в сквален. Изначально, соединение подвергается переносу остатка фосфорной кислоты при помощи фермента АТФ. Продуктом реакции выступает 5′-пирофосфорный эфир. Впрочем, соединение не долговечно. Оно практически моментально трансформируется, в эфир мевалоната. Цепочка последующих преобразований достаточно запутана, поэтому проще ограничиться фактом. Результатом этих процессов становится образование сквалена. Реакция циклизации соединения приводит к формированию ланостерина, с последующим биосинтезом холестерола.
Остается добавить, что преимущественно процесс протекает в печени. Поэтому дисфункции органа, способны вызывать нарушения липидного баланса. При нормальной работе, печень производит ежесуточную норму холестерина, с учетом вещества, поступающего с продуктами. Этот факт еще раз опровергает распространенное заблуждение о вреде пищи с высоким содержанием холестерина. Здоровая печень, способна самостоятельно контролировать уровень вещества.
Окисление липидов в организме – это различные типы реакций, которые имеют как положительные, так и отрицательные последствия для человеческого организма.
Функции липидов разнообразны. Некоторые группы липидов необходимы в качестве, так называемых, строительных «кирпичиков» для биологических мембран
Окисление липидов в организме – это различные типы реакций, которые имеют как положительные, так и отрицательные последствия для человеческого организма.
Функции липидов разнообразны. Некоторые группы липидов необходимы в качестве, так называемых, строительных «кирпичиков» для биологических мембран
Источники:
http://www.xumuk.ru/biochem/304.html
http://vinograd.info/knigi/osnovy-biohimii-vinodeliya/metabolizm-lipidov-i-ih-rol-3.html
http://sosudportal.ru/li/sintez-lipidov-process.html




